

Osteopontin promotes hepatocellular carcinoma progression through inducing JAK2/STAT3/NOX1-mediated ROS production
- PMID: 35418176
- PMCID: PMC9008047
- DOI: 10.1038/s41419-022-04806-9
Osteopontin promotes hepatocellular carcinoma progression through inducing JAK2/STAT3/NOX1-mediated ROS production
Abstract
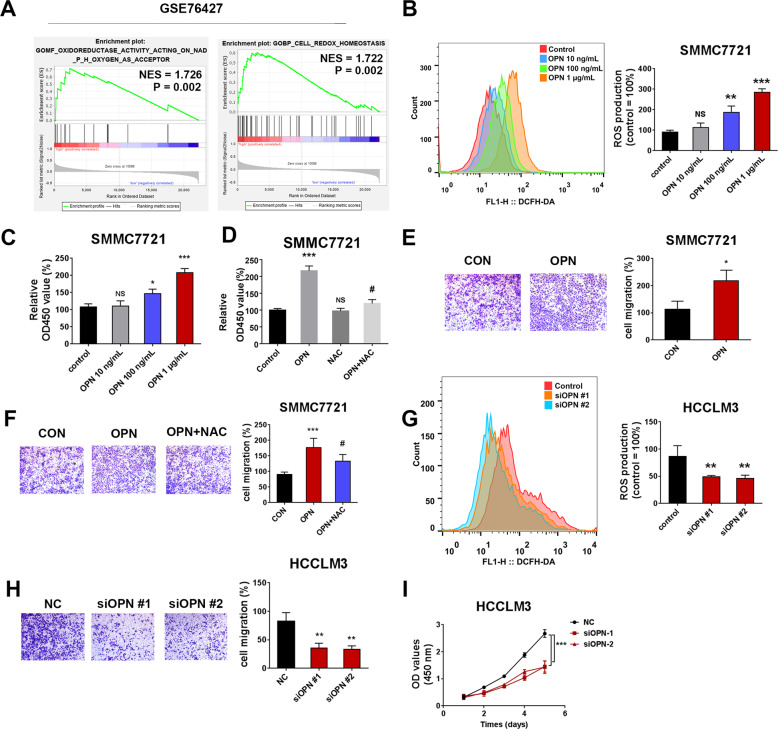
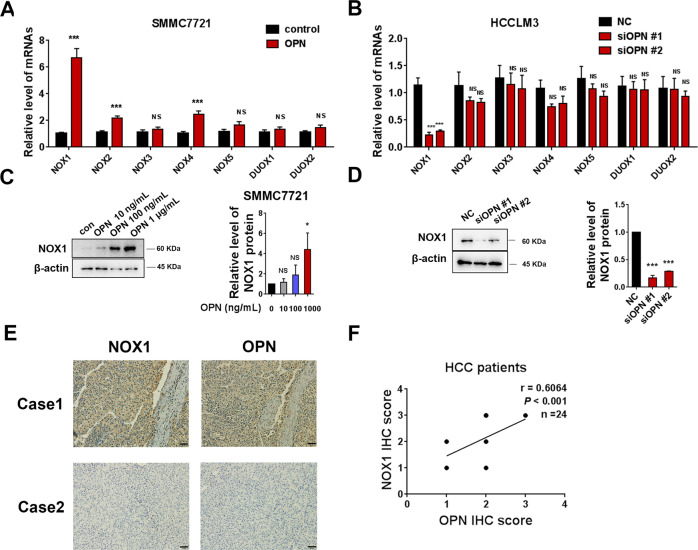
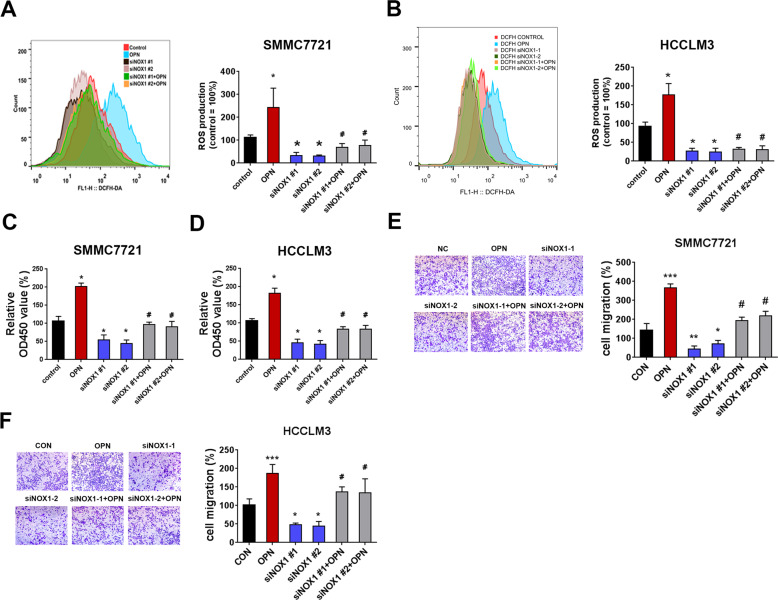
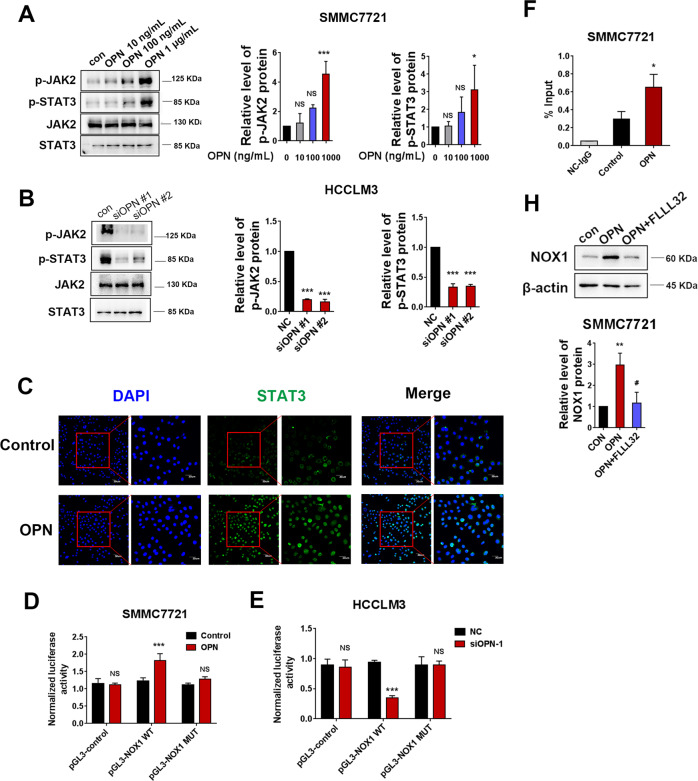
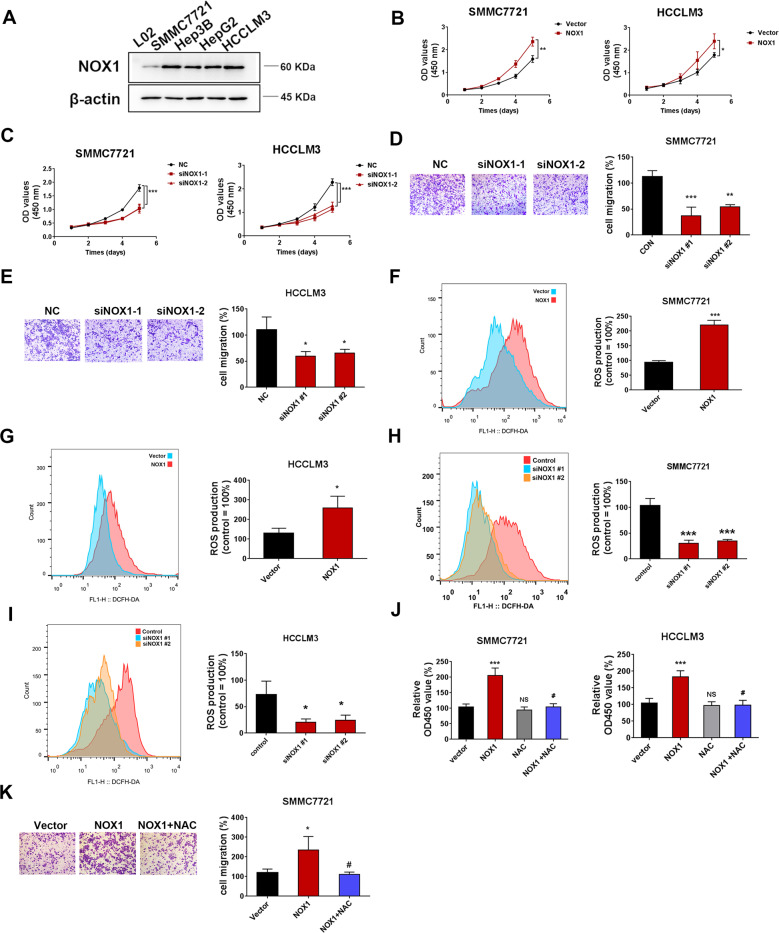
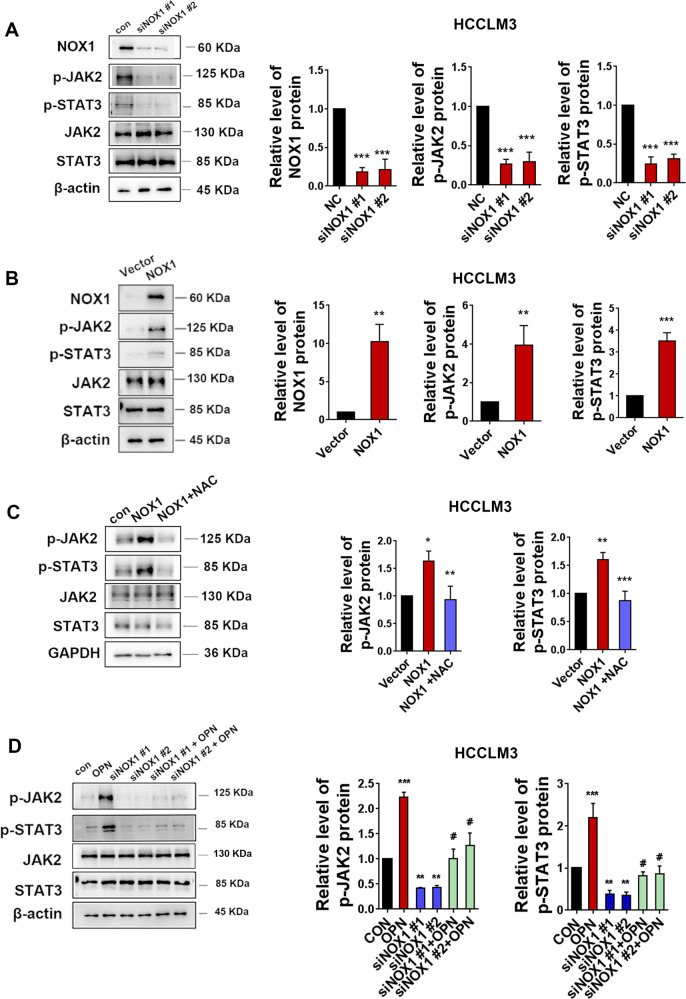
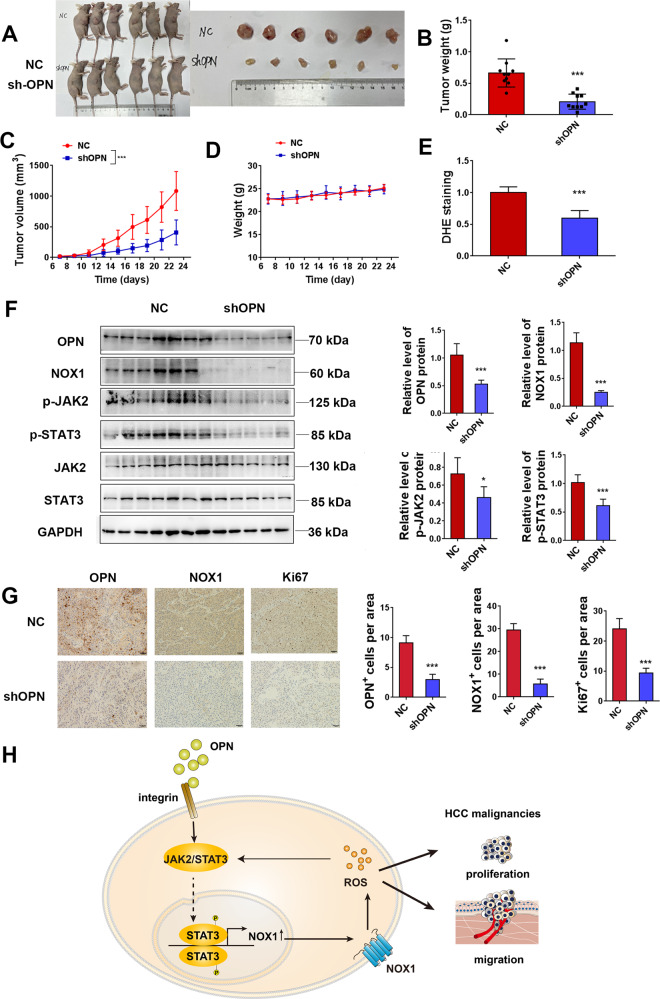
Osteopontin (OPN) is a multifunctional cytokine that can impact cancer progression. Therefore, it is crucial to determine the key factors involved in the biological role of OPN for the development of treatment. Here, we investigated that OPN promoted hepatocellular carcinoma (HCC) cell proliferation and migration by increasing Reactive oxygen species (ROS) production and disclosed the underlying mechanism. Knockdown of OPN suppressed ROS production in vitro and in vivo, whereas treatment with human recombinant OPN produced the opposite effect. N-Acetyl-L-cysteine (NAC, ROS scavenger) partially blocked HCC cell proliferation and migration induced by OPN. Mechanistically, OPN induced ROS production in HCC cells by upregulating the expression of NADPH oxidase 1 (NOX1). NOX1 knockdown in HCC cells partially abrogated the cell proliferation and migration induced by OPN. Moreover, inhibition of JAK2/STAT3 phosphorylation effectively decreased the transcription of NOX1, upregulated by OPN. In addition, NOX1 overexpression increased JAK2 and STAT3 phosphorylation by increasing ROS production, creating a positive feedback loop for stimulating JAK2/STAT3 signaling induced by OPN. This study for the first time demonstrated that HCC cells utilized OPN to generate ROS for tumor progression, and disruption of OPN/NOX1 axis might be a promising therapeutic strategy for HCC.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
