

Nonlinear control of transcription through enhancer-promoter interactions
- PMID: 35418676
- PMCID: PMC9021019
- DOI: 10.1038/s41586-022-04570-y
Nonlinear control of transcription through enhancer-promoter interactions
Abstract
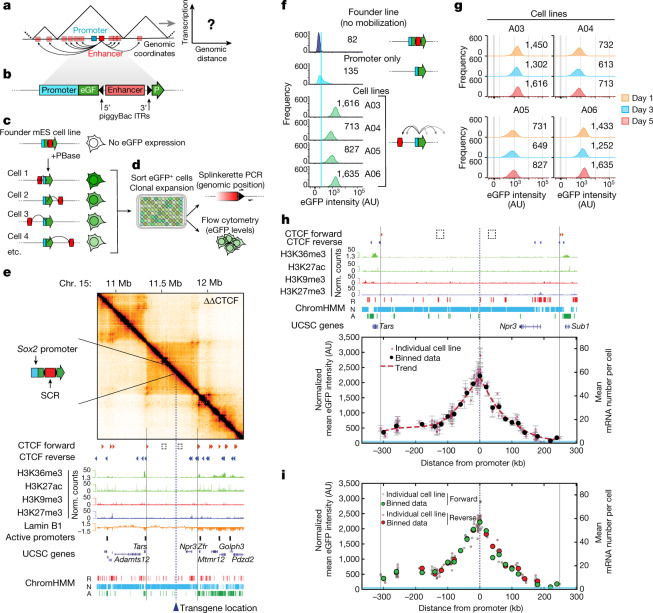
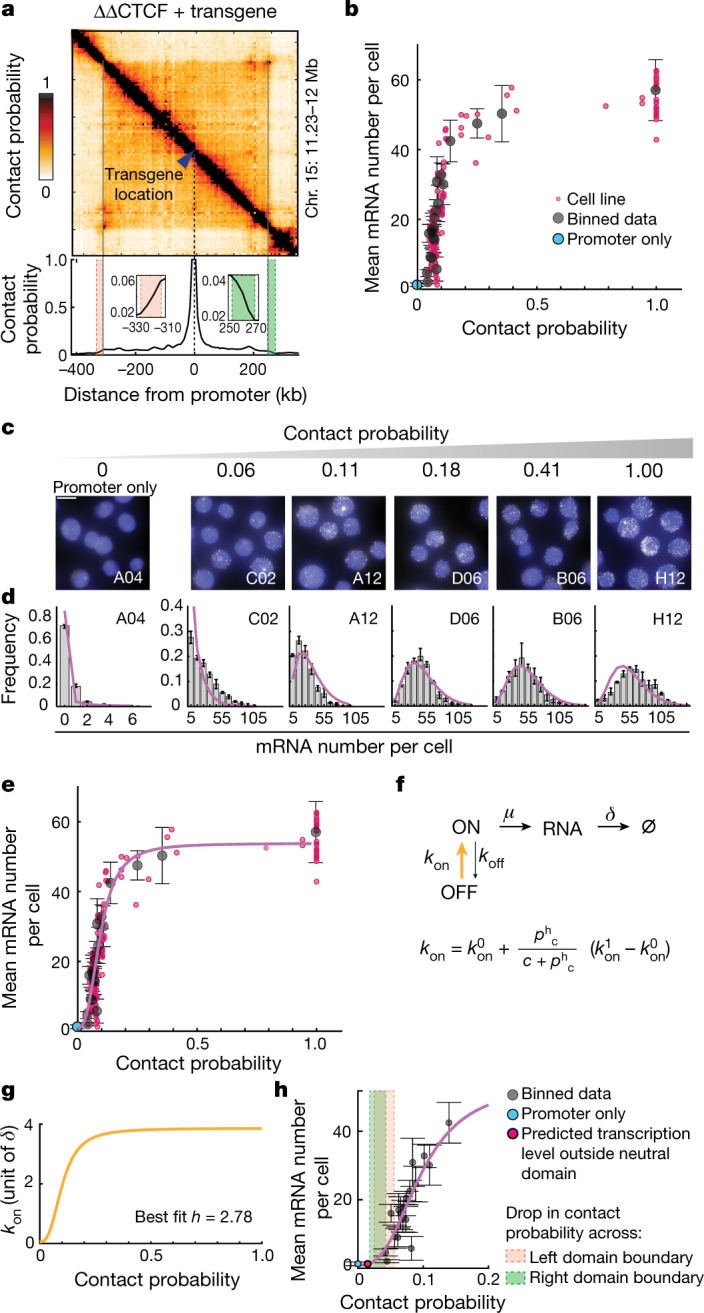
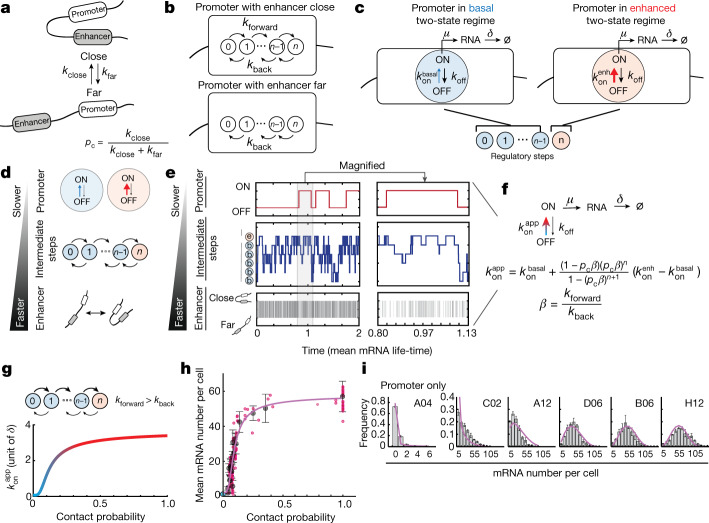
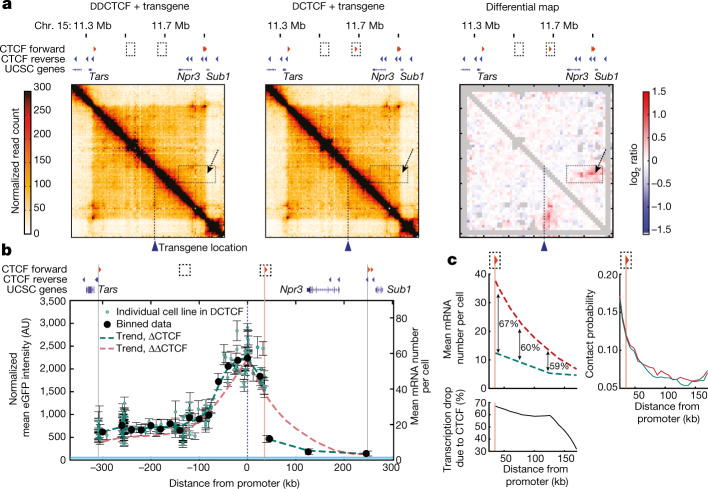
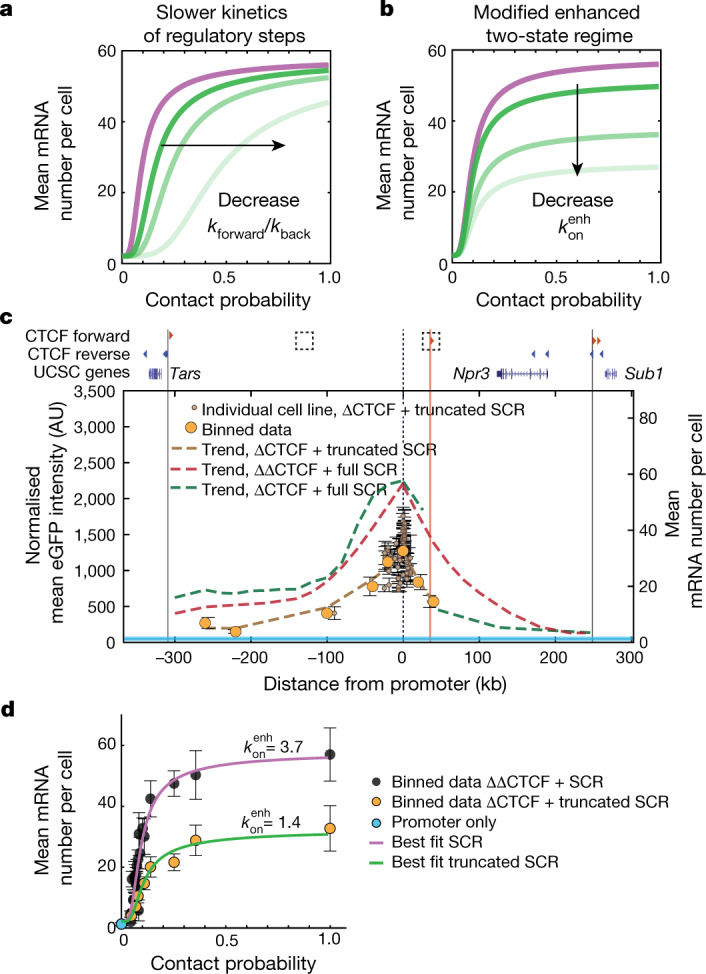
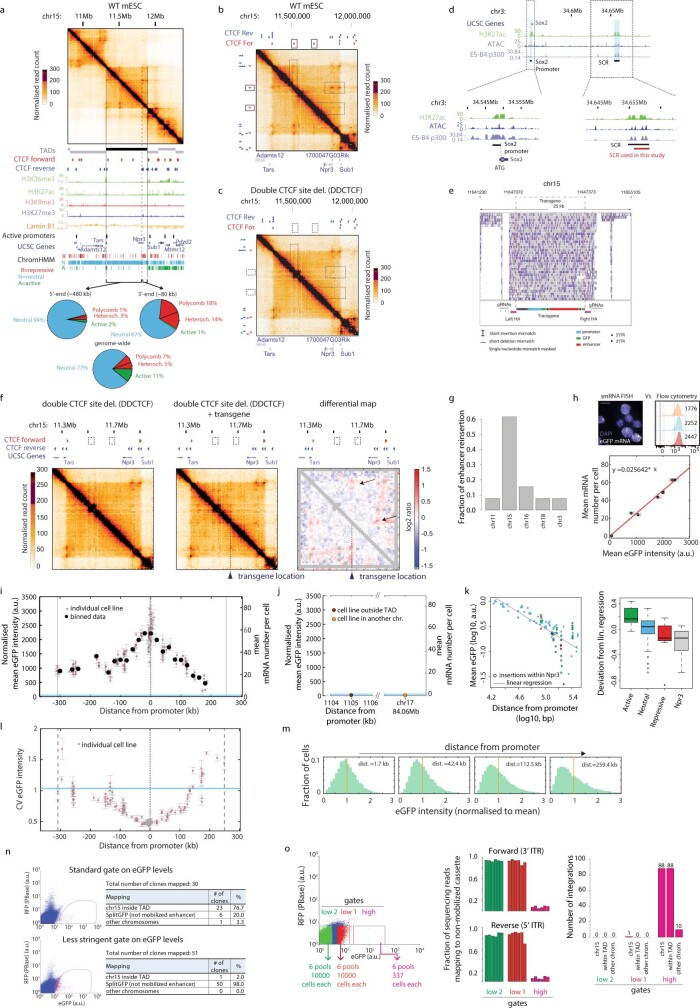
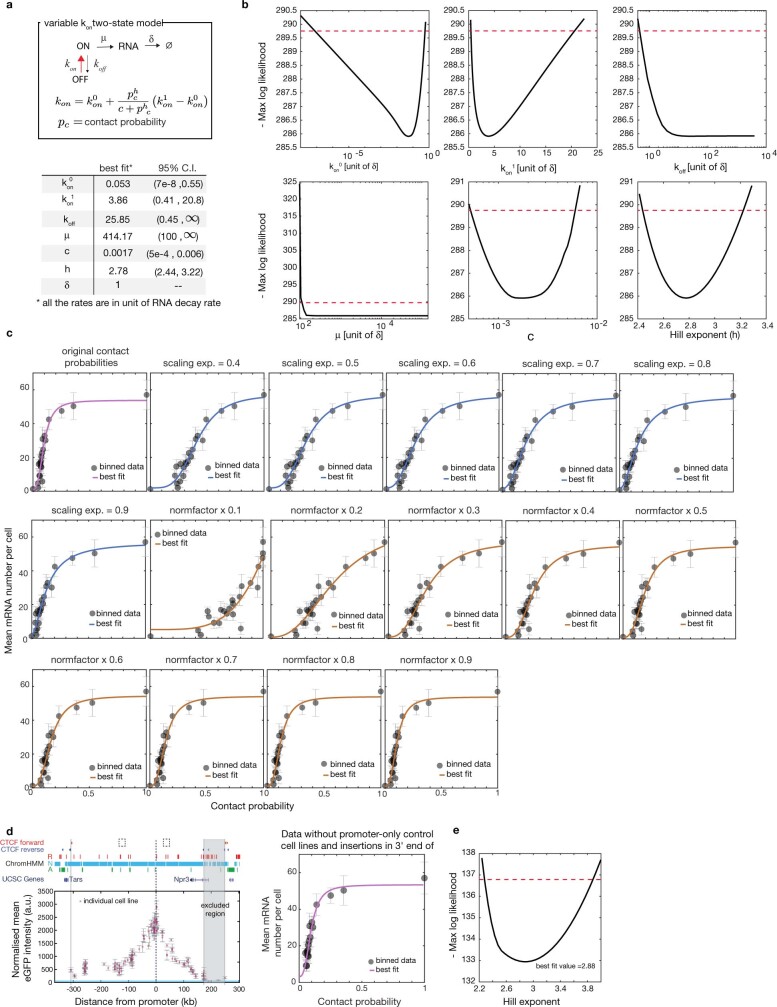
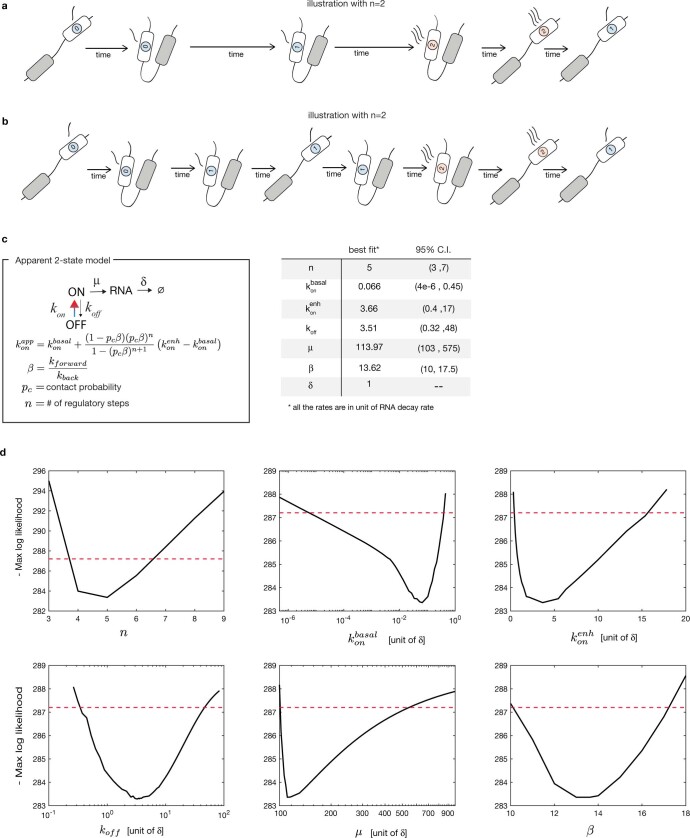
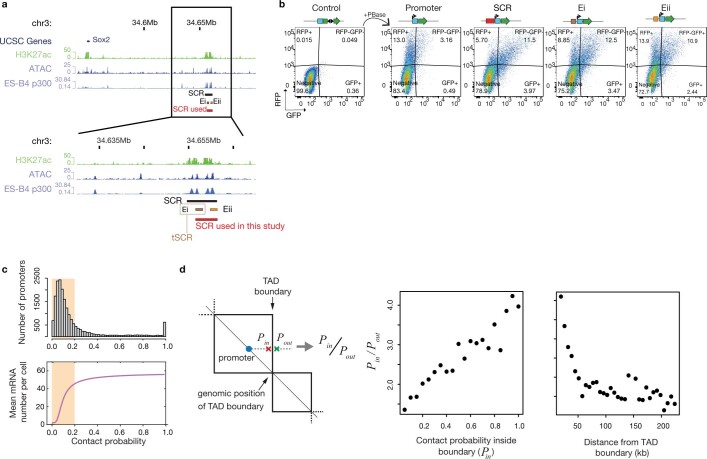
Chromosome structure in mammals is thought to regulate transcription by modulating three-dimensional interactions between enhancers and promoters, notably through CTCF-mediated loops and topologically associating domains (TADs)1-4. However, how chromosome interactions are actually translated into transcriptional outputs remains unclear. Here, to address this question, we use an assay to position an enhancer at large numbers of densely spaced chromosomal locations relative to a fixed promoter, and measure promoter output and interactions within a genomic region with minimal regulatory and structural complexity. A quantitative analysis of hundreds of cell lines reveals that the transcriptional effect of an enhancer depends on its contact probabilities with the promoter through a nonlinear relationship. Mathematical modelling suggests that nonlinearity might arise from transient enhancer-promoter interactions being translated into slower promoter bursting dynamics in individual cells, therefore uncoupling the temporal dynamics of interactions from those of transcription. This uncovers a potential mechanism of how distal enhancers act from large genomic distances, and of how topologically associating domain boundaries block distal enhancers. Finally, we show that enhancer strength also determines absolute transcription levels as well as the sensitivity of a promoter to CTCF-mediated transcriptional insulation. Our measurements establish general principles for the context-dependent role of chromosome structure in long-range transcriptional regulation.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Comment in
-
Brief encounters: The relationship between enhancer proximity and gene expression.Dev Cell. 2022 Jun 20;57(12):1439-1441. doi: 10.1016/j.devcel.2022.05.020. Dev Cell. 2022. PMID: 35728521
-
Enhancer-promoter communication: unraveling enhancer strength and positioning within a given topologically associating domain (TAD).Signal Transduct Target Ther. 2022 Aug 12;7(1):281. doi: 10.1038/s41392-022-01114-8. Signal Transduct Target Ther. 2022. PMID: 35961952 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
