

Carrier Proteins Facilitate the Generation of Antipolysaccharide Immunity via Multiple Mechanisms
- PMID: 35420488
- PMCID: PMC9239039
- DOI: 10.1128/mbio.03790-21
Carrier Proteins Facilitate the Generation of Antipolysaccharide Immunity via Multiple Mechanisms
Abstract
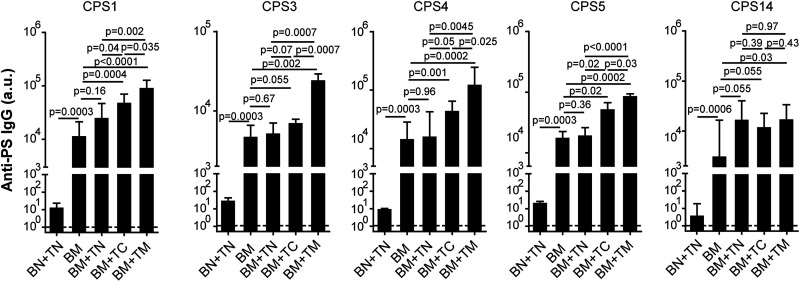
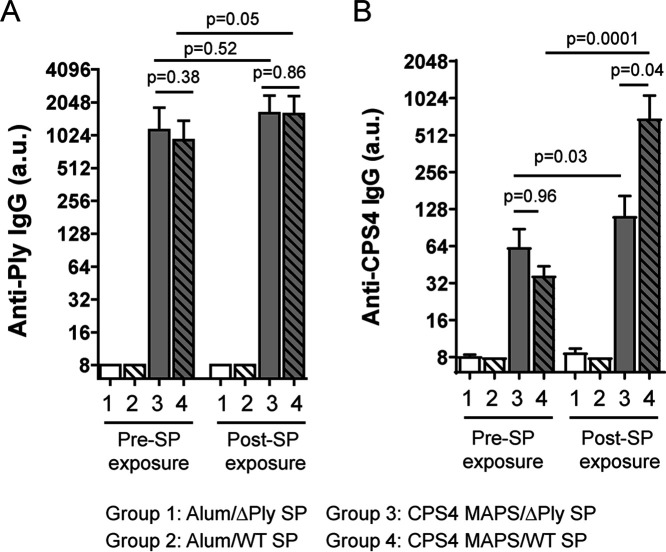
Capsular polysaccharides (CPSs) are important antigenic targets against bacterial infections. As T-independent antigens, however, CPSs elicit short-lived immune responses in adults and are poorly immunogenic in young children. Coupling CPS with protein carriers enhances anti-CPS responses and generates long-lasting immune memory. However, the mechanisms whereby carrier proteins accomplish this are not fully understood. Here, we dissect different mechanisms whereby carrier proteins enhance anti-CPS immunity. We show how coupling CPS with protein carriers modifies the interaction of CPS with antigen-presenting cells, enables a dual-activation mechanism for CPS-specific B cells via interaction with CPS- or carrier-specific T helper cells, and potentiates the recall of anti-CPS responses by engaging memory T helper cells during subsequent vaccination or bacterial exposure. Our findings provide new insights into the immunological basis of carrier-mediated anti-CPS immunity and may help in the design of more effective CPS-based vaccines. IMPORTANCE Polysaccharide capsules, the outermost shells of many bacterial pathogens, play a role in pathogenesis and protect bacteria against the immune system. Generating antipolysaccharide antibodies by vaccination has provided effective protection against infectious diseases caused by encapsulated bacteria. However, most pure polysaccharide preparations are poorly immunogenic, particularly in young children. To circumvent this problem, vaccines have been developed using polysaccharides associated with protein carriers. The precise mechanism whereby protein carriers enhance the immunogenicity of the polysaccharide remains unclear. The significance of our research is in elucidating the different roles played by carriers in facilitating polysaccharide processing and presentation, priming polysaccharide-specific B cells, and potentiating recall antipolysaccharide responses. Overall, our work provides new insights into the immunological basis of carrier-mediated antipolysaccharide immunity and may help in the design of more effective polysaccharide-based vaccines.
Keywords: capsular polysaccharide; immune mechanisms; immunization.
Conflict of interest statement
The authors declare a conflict of interest. F.Z., R.M., and Y.-J.L. are named inventors on filed patents related to the MAPS technology and are scientific founders, consultants, and equity owners at Affinivax, a biotechnology company based in Cambridge, MA, that is devoted to the development of vaccines for developing and developed countries, based on the MAPS technology. R.M. is also a part-time employee at Affinivax, as Chief Scientific Officer. Affinivax had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
F.Z., R.M., and Y.-J.L. are inventors on filed patents related to the MAPS technology and are scientific founders, consultants, and equity owners at Affinivax, a biotechnology company using the MAPS technology for vaccine development. R.M. is also Chief Scientific Officer of Affinivax. Affinivax had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Figures
Similar articles
-
Development of meningococcal polysaccharide conjugate vaccine that can elicit long-lasting and strong cellular immune response with hepatitis B core antigen virus-like particles as a novel carrier protein.Vaccine. 2019 Feb 8;37(7):956-964. doi: 10.1016/j.vaccine.2018.12.073. Epub 2019 Jan 14. Vaccine. 2019. PMID: 30655174
-
A Structural Model for the Ligand Binding of Pneumococcal Serotype 3 Capsular Polysaccharide-Specific Protective Antibodies.mBio. 2021 Jun 29;12(3):e0080021. doi: 10.1128/mBio.00800-21. Epub 2021 Jun 1. mBio. 2021. PMID: 34061603 Free PMC article.
-
Vaccines against polysaccharide antigens.Curr Drug Targets Infect Disord. 2001 Nov;1(3):325-34. doi: 10.2174/1568005014605964. Curr Drug Targets Infect Disord. 2001. PMID: 12455405 Review.
-
Preparation and preclinical evaluation of experimental group B streptococcus type III polysaccharide-cholera toxin B subunit conjugate vaccine for intranasal immunization.Vaccine. 2000 Nov 22;19(7-8):850-61. doi: 10.1016/s0264-410x(00)00226-7. Vaccine. 2000. PMID: 11115709
-
Differential regulation of polysaccharide-specific antibody responses to isolated polysaccharides, conjugate vaccines, and intact Gram-positive versus Gram-negative extracellular bacteria.Vaccine. 2016 Jun 24;34(30):3542-8. doi: 10.1016/j.vaccine.2015.12.077. Epub 2016 Apr 29. Vaccine. 2016. PMID: 27133879 Review.
Cited by
-
Novel Klebsiella pneumoniae and Pseudomonas aeruginosa MAPS vaccine combining O polysaccharides and pathogen-specific proteins.mBio. 2025 Aug 13;16(8):e0080725. doi: 10.1128/mbio.00807-25. Epub 2025 Jun 26. mBio. 2025. PMID: 40569061 Free PMC article.
-
Preclinical Immunogenicity and Efficacy of a Multiple Antigen-Presenting System (MAPSTM) SARS-CoV-2 Vaccine.Vaccines (Basel). 2022 Jul 3;10(7):1069. doi: 10.3390/vaccines10071069. Vaccines (Basel). 2022. PMID: 35891233 Free PMC article.
-
A Bivalent MAPS Vaccine Induces Protective Antibody Responses against Salmonella Typhi and Paratyphi A.Vaccines (Basel). 2022 Dec 30;11(1):91. doi: 10.3390/vaccines11010091. Vaccines (Basel). 2022. PMID: 36679935 Free PMC article.
-
Conjugation Mechanism for Pneumococcal Glycoconjugate Vaccines: Classic and Emerging Methods.Bioengineering (Basel). 2022 Dec 6;9(12):774. doi: 10.3390/bioengineering9120774. Bioengineering (Basel). 2022. PMID: 36550980 Free PMC article. Review.
-
Epidemiological and clinical characteristics of pertussis in children and their close contacts in households: A cross-sectional survey in Zhejiang Province, China.Front Pediatr. 2022 Aug 18;10:976796. doi: 10.3389/fped.2022.976796. eCollection 2022. Front Pediatr. 2022. PMID: 36061393 Free PMC article.
References
-
- Wen Z, Zhang J-R. 2015. Bacterial capsules, p 33–53. In Tang Y-W, Sussman M, Liu D, Poxton I, Schwartzman J (ed), Molecular medical microbiology, 2nd ed. Academic Press, Boston, MA. 10.1016/B978-0-12-397169-2.00003-2. - DOI
-
- Mitsi E, Roche AM, Reiné J, Zangari T, Owugha JT, Pennington SH, Gritzfeld JF, Wright AD, Collins AM, van Selm S, de Jonge MI, Gordon SB, Weiser JN, Ferreira DM. 2017. Agglutination by anti-capsular polysaccharide antibody is associated with protection against experimental human pneumococcal carriage. Mucosal Immunol 10:385–394. doi:10.1038/mi.2016.71. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
