

Myoblast deactivation within engineered human skeletal muscle creates a transcriptionally heterogeneous population of quiescent satellite-like cells
- PMID: 35421801
- PMCID: PMC9289780
- DOI: 10.1016/j.biomaterials.2022.121508
Myoblast deactivation within engineered human skeletal muscle creates a transcriptionally heterogeneous population of quiescent satellite-like cells
Abstract
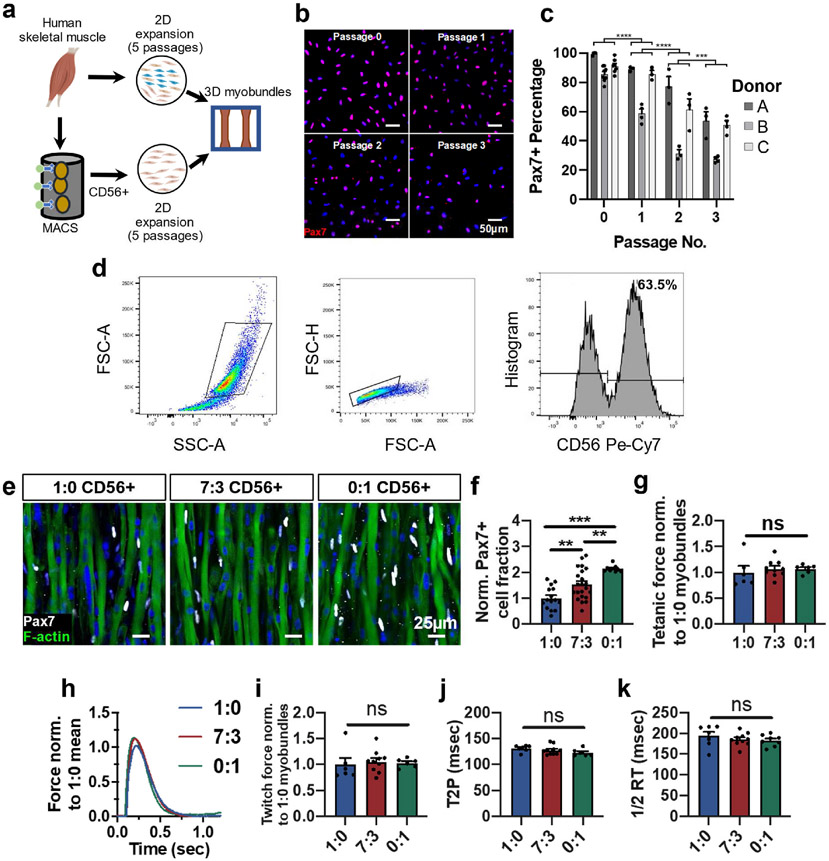
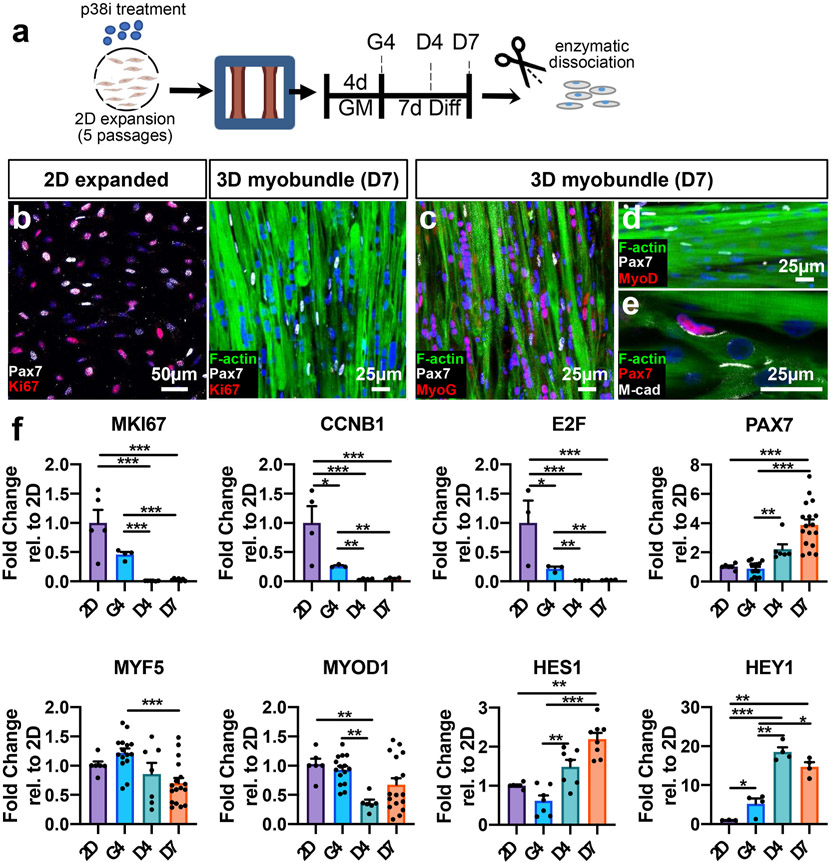
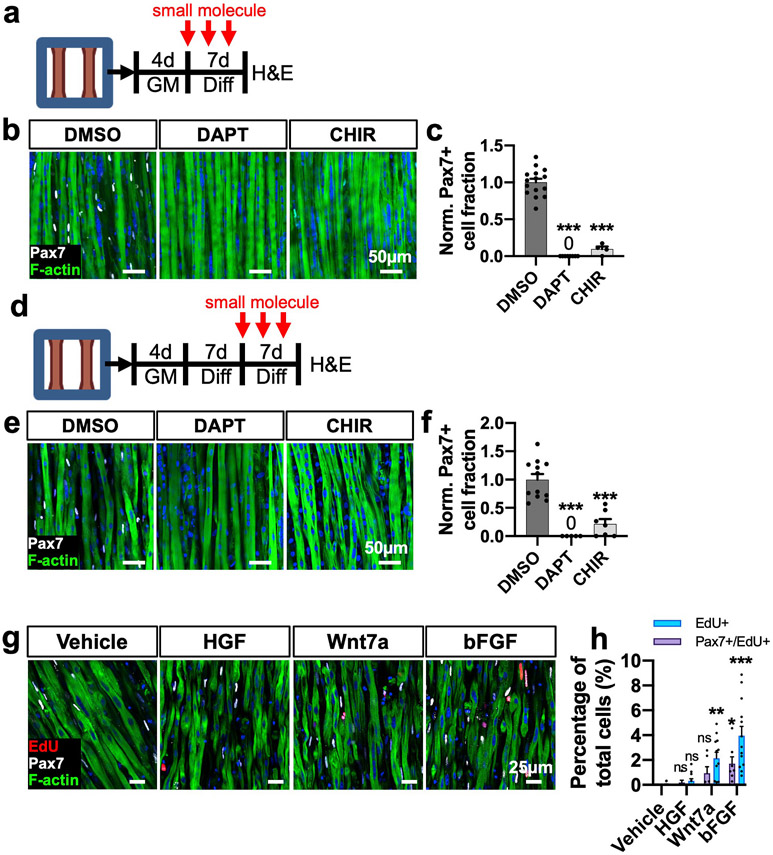
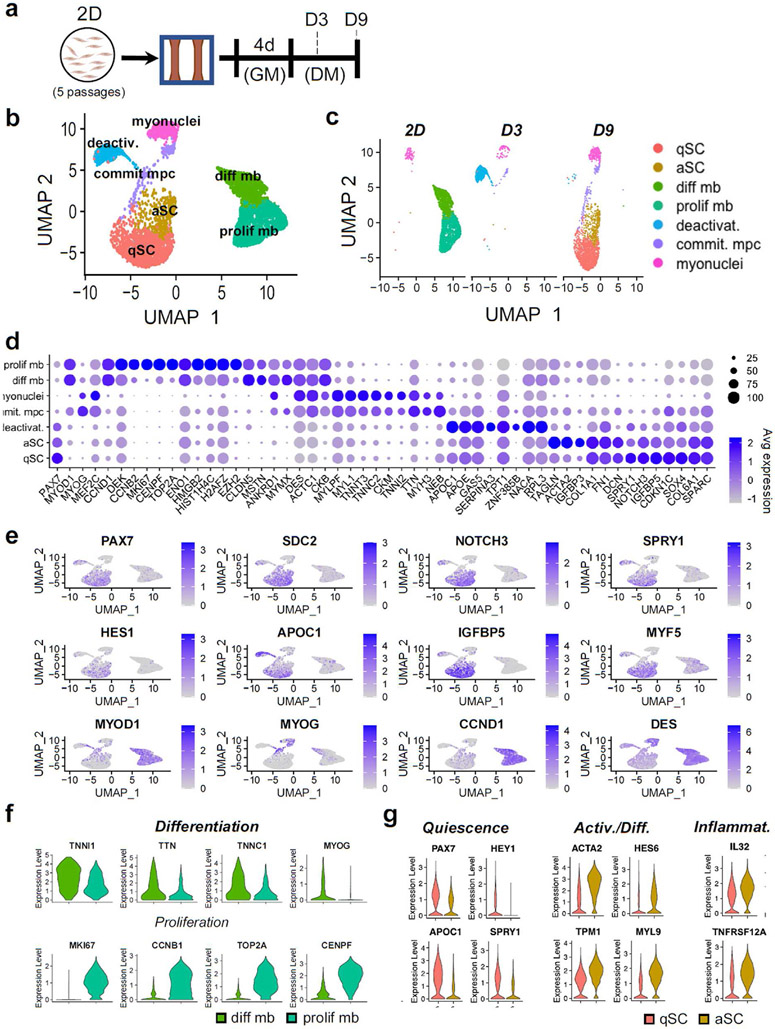
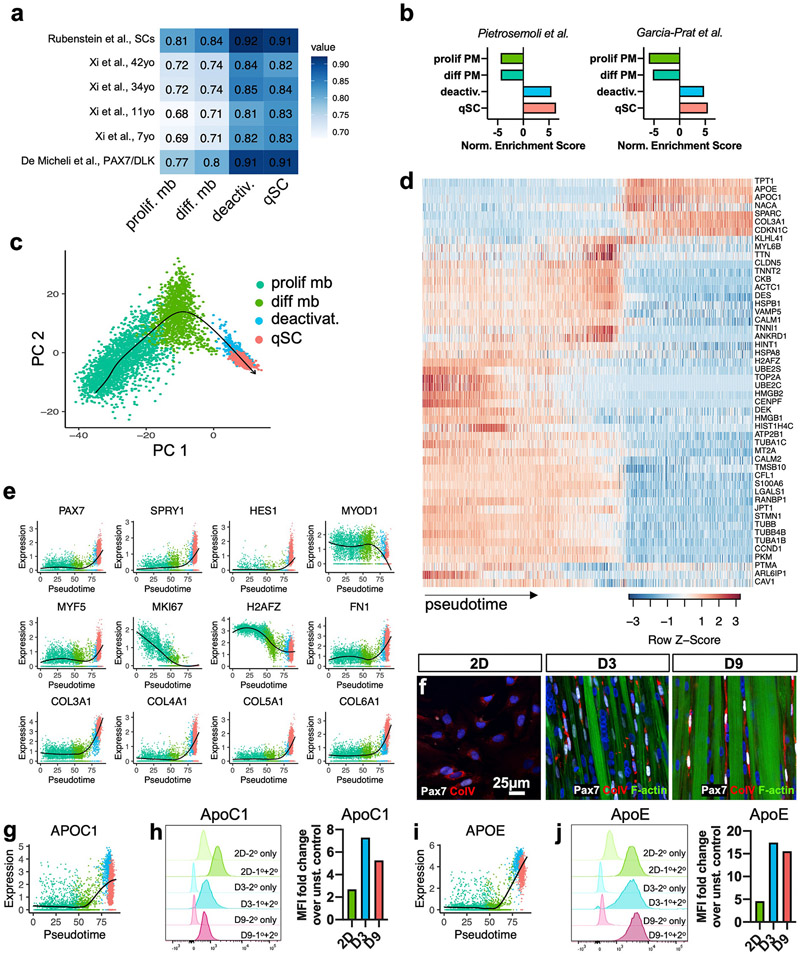
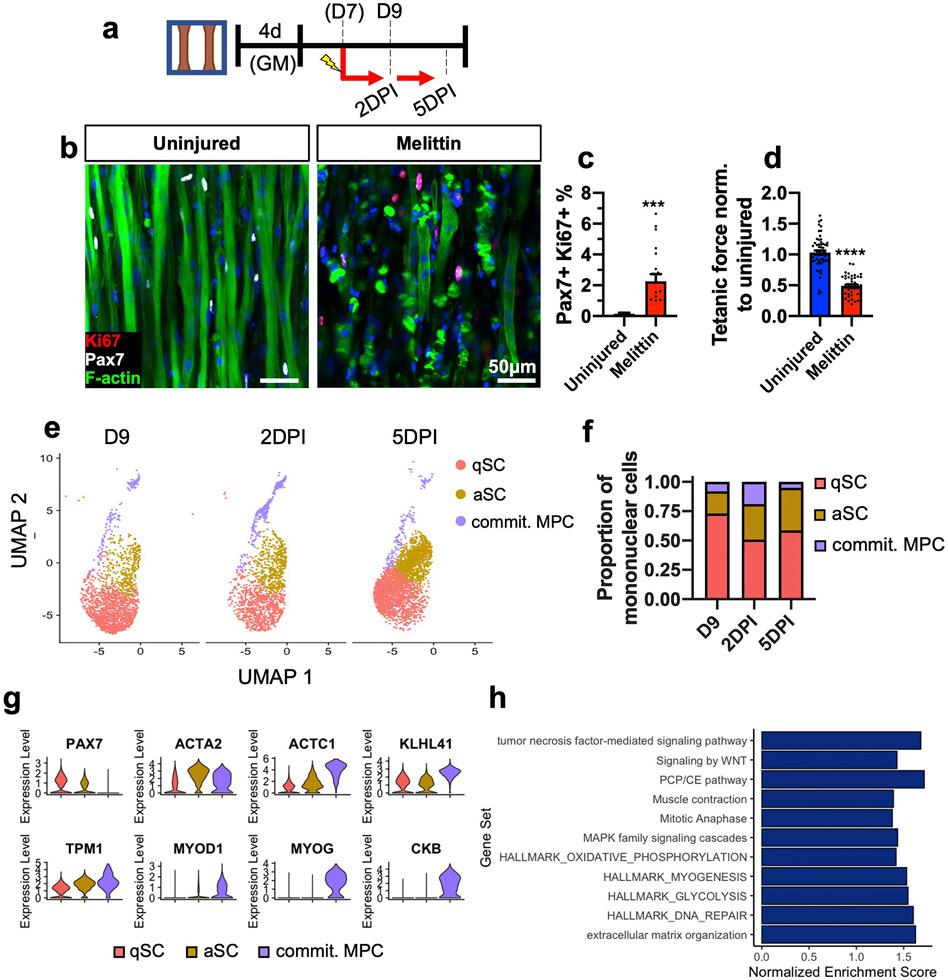
Satellite cells (SCs), the adult Pax7-expressing stem cells of skeletal muscle, are essential for muscle repair. However, in vitro investigations of SC function are challenging due to isolation-induced SC activation, loss of native quiescent state, and differentiation to myoblasts. In the present study, we optimized methods to deactivate in vitro expanded human myoblasts within a 3D culture environment of engineered human skeletal muscle tissues ("myobundles"). Immunostaining and gene expression analyses revealed that a fraction of myoblasts within myobundles adopted a quiescent phenotype (3D-SCs) characterized by increased Pax7 expression, cell cycle exit, and activation of Notch signaling. Similar to native SCs, 3D-SC quiescence is regulated by Notch and Wnt signaling while loss of quiescence and reactivation of 3D-SCs can be induced by growth factors including bFGF. Myobundle injury with a bee toxin, melittin, induces robust myofiber fragmentation, functional decline, and 3D-SC proliferation. By applying single cell RNA-sequencing (scRNA-seq), we discover the existence of two 3D-SC subpopulations (quiescent and activated), identify deactivation-associated gene signature using trajectory inference between 2D myoblasts and 3D-SCs, and characterize the transcriptomic changes within reactivated 3D-SCs in response to melittin-induced injury. These results demonstrate the ability of an in vitro engineered 3D human skeletal muscle environment to support the formation of a quiescent and heterogeneous SC population recapitulating several aspects of the native SC phenotype, and provide a platform for future studies of human muscle regeneration and disease-associated SC dysfunction.
Keywords: Activation; Engineered skeletal muscle; Human; Quiescence; Satellite cells; scRNA-seq.
Copyright © 2022 Elsevier Ltd. All rights reserved.
Figures
References
-
- Mourikis P, Sambasivan R, Castel D, Rocheteau P, Bizzarro V, Tajbakhsh S, A critical requirement for notch signaling in maintenance of the quiescent skeletal muscle stem cell state, Stem Cell. 30 (2) (2012) 243–252. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
