

Adaptive responses of marine diatoms to zinc scarcity and ecological implications
- PMID: 35422102
- PMCID: PMC9010474
- DOI: 10.1038/s41467-022-29603-y
Adaptive responses of marine diatoms to zinc scarcity and ecological implications
Abstract
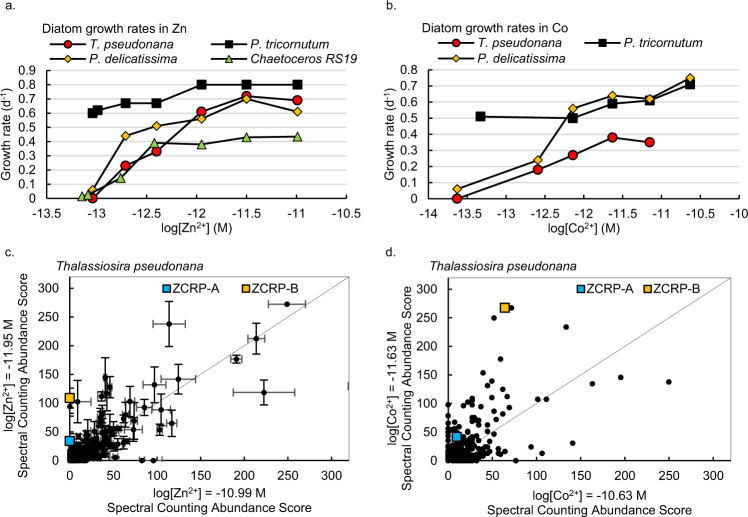
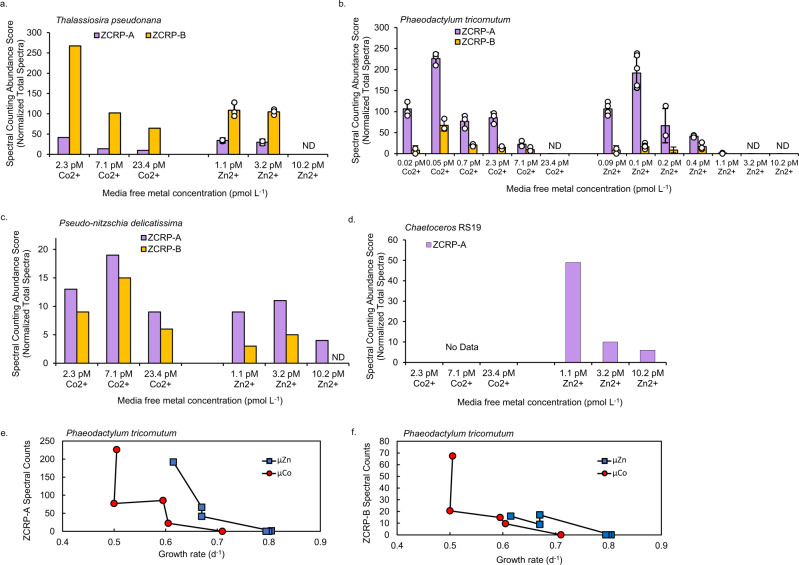
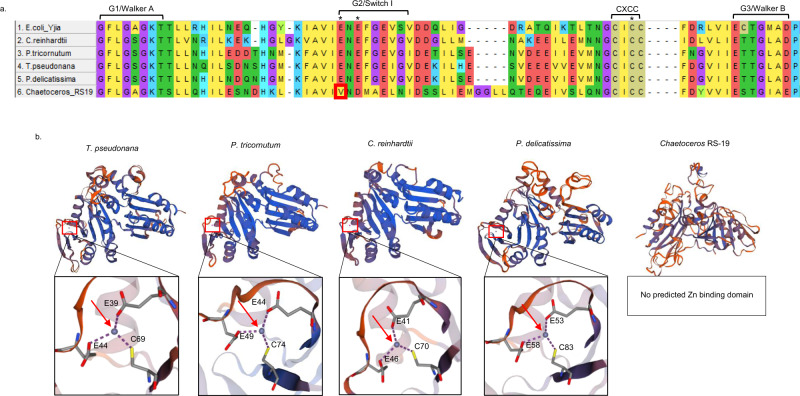
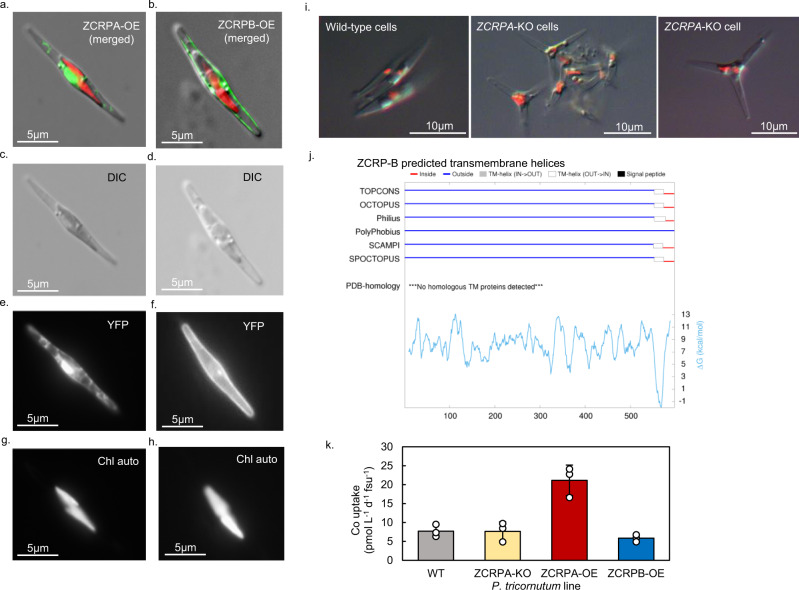
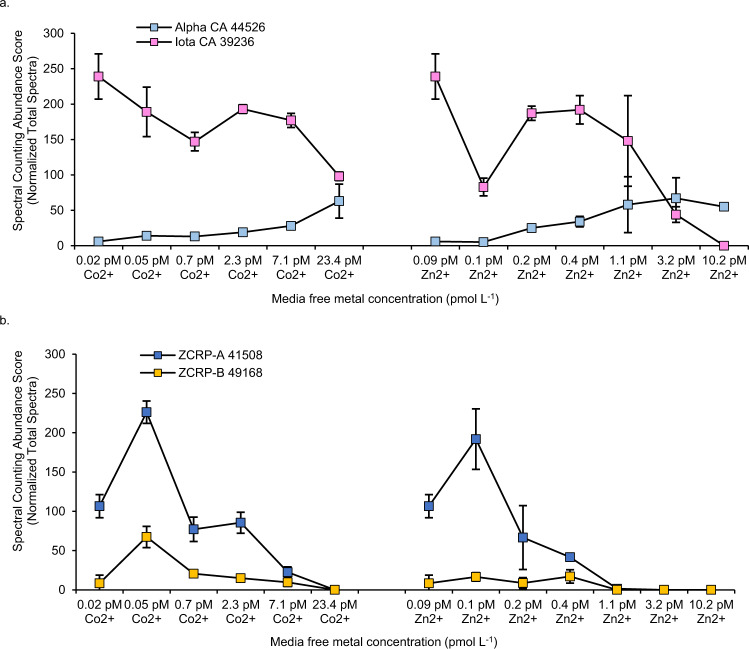
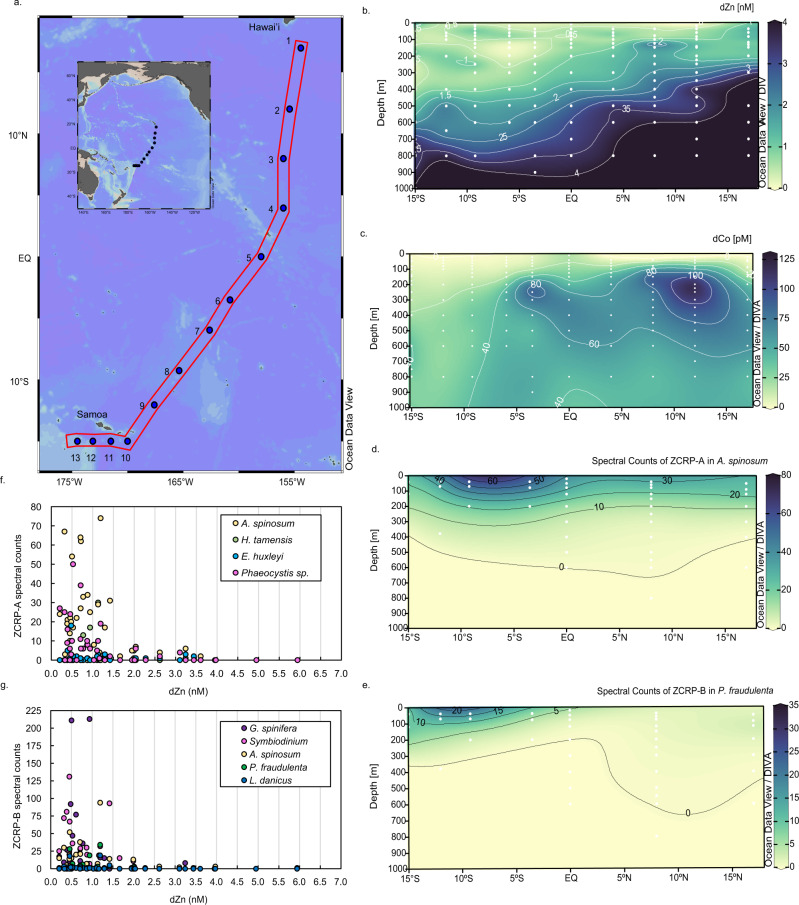
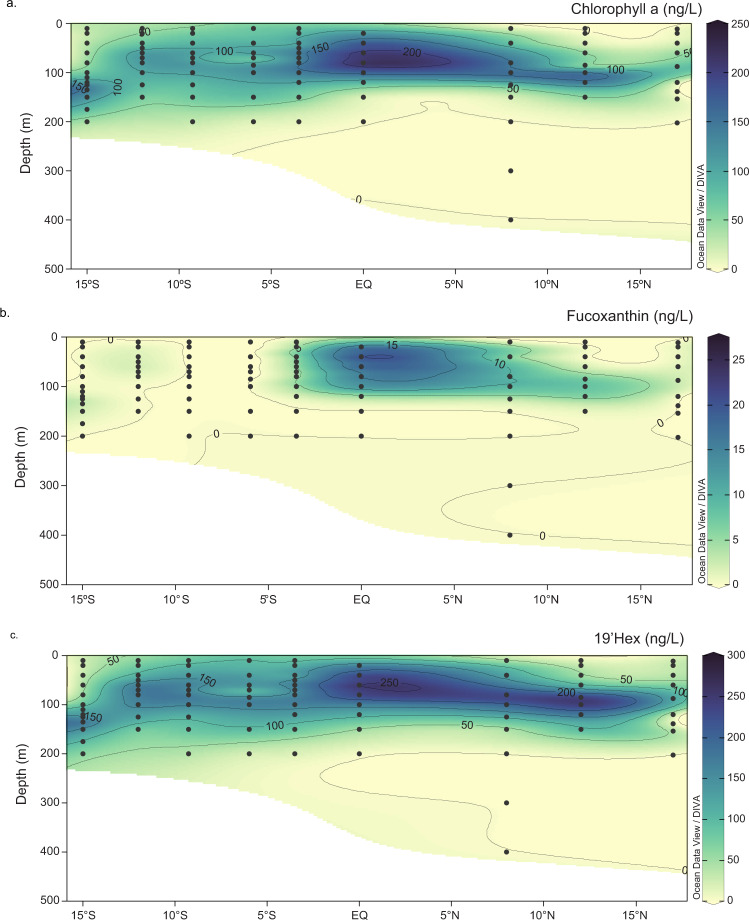
Scarce dissolved surface ocean concentrations of the essential algal micronutrient zinc suggest that Zn may influence the growth of phytoplankton such as diatoms, which are major contributors to marine primary productivity. However, the specific mechanisms by which diatoms acclimate to Zn deficiency are poorly understood. Using global proteomic analysis, we identified two proteins (ZCRP-A/B, Zn/Co Responsive Protein A/B) among four diatom species that became abundant under Zn/Co limitation. Characterization using reverse genetic techniques and homology data suggests putative Zn/Co chaperone and membrane-bound transport complex component roles for ZCRP-A (a COG0523 domain protein) and ZCRP-B, respectively. Metaproteomic detection of ZCRPs along a Pacific Ocean transect revealed increased abundances at the surface (<200 m) where dZn and dCo were scarcest, implying Zn nutritional stress in marine algae is more prevalent than previously recognized. These results demonstrate multiple adaptive responses to Zn scarcity in marine diatoms that are deployed in low Zn regions of the Pacific Ocean.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Sunda WG, Huntsman SA. Cobalt and zinc interreplacement in marine phytoplankton: Biological and geochemical implications. Limnol. Oceanogr. 1995;40:1404–1417. doi: 10.4319/lo.1995.40.8.1404. - DOI
-
- Bruland KW. Oceanographic distributions of cadmium, zinc, nickel, and copper in the North Pacific. Earth Planet. Sci. Lett. 1980;47:176–198. doi: 10.1016/0012-821X(80)90035-7. - DOI
-
- Brand LE, Sunda WG, Guillard RRL. Limitation of marine phytoplankton reproductive rates by zinc, manganese, and iron. Limnol. Oceanogr. 1983;28:1182–1198. doi: 10.4319/lo.1983.28.6.1182. - DOI
-
- Morel FMM, et al. Zinc and carbon co-limitation of marine phytoplankton. Nature. 1994;369:740–742. doi: 10.1038/369740a0. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
