

Engineering bacteria to control electron transport altering the synthesis of non-native polymer
- PMID: 35424487
- PMCID: PMC8978702
- DOI: 10.1039/d1ra06403g
Engineering bacteria to control electron transport altering the synthesis of non-native polymer
Abstract
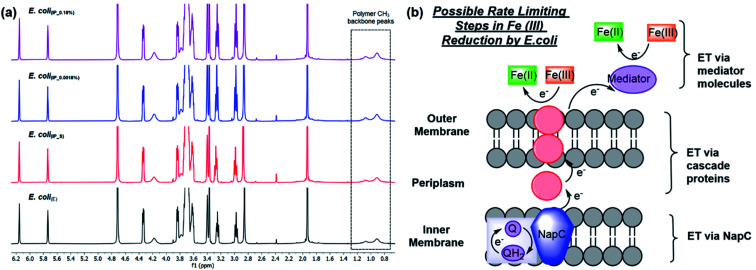
The use of bacteria as catalysts for radical polymerisations of synthetic monomers has recently been established. However, the role of trans Plasma Membrane Electron Transport (tPMET) in modulating these processes is not well understood. We sort to study this by genetic engineering a part of the tPMET system NapC in E. coli. We show that this engineering altered the rate of extracellular electron transfer coincided with an effect on cell-mediated polymerisation using a model monomer. A plasmid with arabinose inducible PBAD promoters were shown to upregulate NapC protein upon induction at total arabinose concentrations of 0.0018% and 0.18%. These clones (E. coli (IP_0.0018%) and E. coli (IP_0.18%), respectively) were used in iron-mediated atom transfer radical polymerisation (Fe ATRP), affecting the nature of the polymerisation, than cultures containing suppressed or empty plasmids (E. coli (IP_S) and E. coli (E), respectively). These results lead to the hypothesis that EET (Extracellular Electron Transfer) in part modulates cell instructed polymerisations.
This journal is © The Royal Society of Chemistry.
Conflict of interest statement
There are no conflicts to declare.
Figures


References
-
- Sherman H. G. Jovanovic C. Abuawad A. Kim D.-H. Collins H. Dixon J. E. Cavanagh R. Markus R. Stolnik S. Rawson F. J. Mechanistic insight into heterogeneity of trans-plasma membrane electron transport in cancer cell types. Biochim. Biophys. Acta, Bioenerg. 2019;1860(8):628–639. doi: 10.1016/j.bbabio.2019.06.012. - DOI - PubMed
-
- Robinson A. J. Jain A. Sherman H. G. Hague R. J. Rahman R. Sanjuan-Alberte P. Rawson F. J. Toward Hijacking Bioelectricity in Cancer to Develop New Bioelectronic Medicine. Adv. Ther. 2021;4(3):2000248. doi: 10.1002/adtp.202000248. - DOI
LinkOut - more resources
Full Text Sources