

An integrated systems-level model of ochratoxin A toxicity in the zebrafish (Danio rerio) embryo based on NMR metabolic profiling
- PMID: 35428752
- PMCID: PMC9012740
- DOI: 10.1038/s41598-022-09726-4
An integrated systems-level model of ochratoxin A toxicity in the zebrafish (Danio rerio) embryo based on NMR metabolic profiling
Abstract
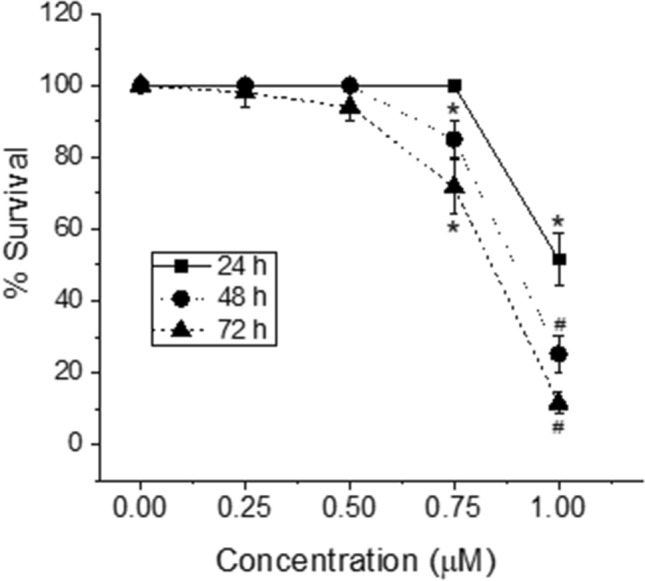
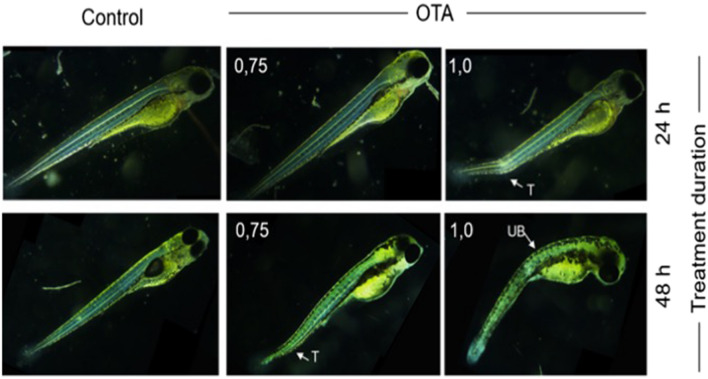
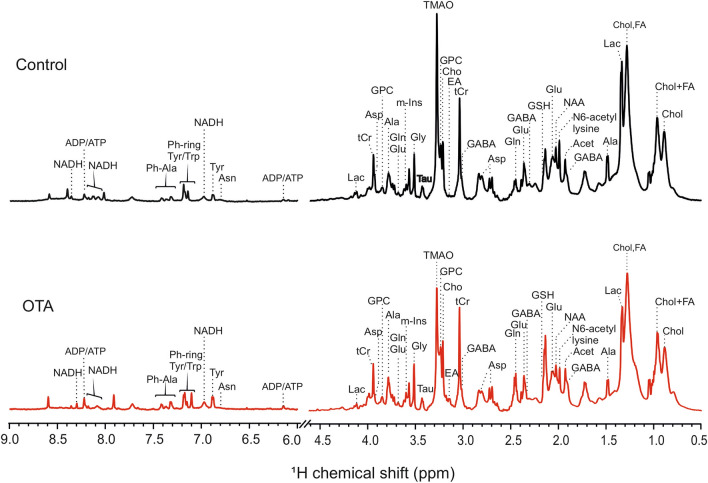
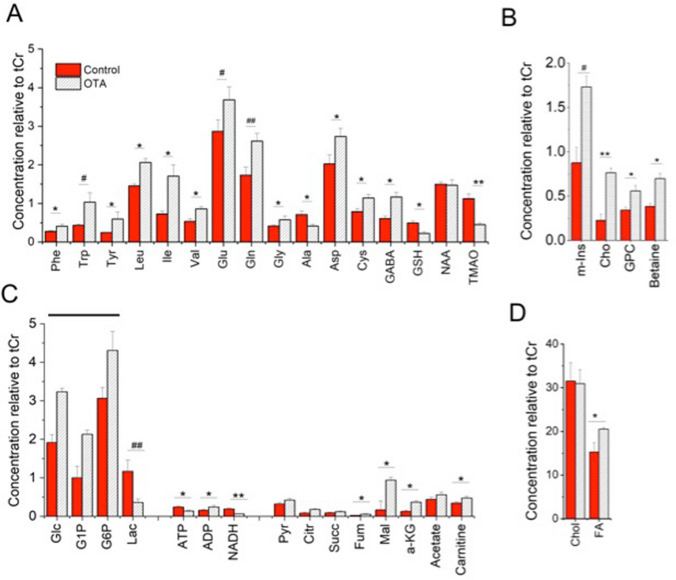
Ochratoxin A (OTA) is one of the most widespread mycotoxin contaminants of agricultural crops. Despite being associated with a range of adverse health effects, a comprehensive systems-level mechanistic understanding of the toxicity of OTA remains elusive. In the present study, metabolic profiling by high-resolution magic angle spinning (HRMAS) NMR, coupled to intact zebrafish embryos, was employed to identify metabolic pathways in relation to a systems-level model of OTA toxicity. Embryotoxicity was observed at sub-micromolar exposure concentrations of OTA. Localization of OTA, based on intrinsic fluorescence, as well as a co-localization of increased reactive oxygen species production, was observed in the liver kidney, brain and intestine of embryos. Moreover, HRMAS NMR showed significant alteration of metabolites related to targeting of the liver (i.e., hepatotoxicity), and pathways associated with detoxification and oxidative stress, and mitochondrial energy metabolism. Based on metabolic profiles, and complementary assays, an integrated model of OTA toxicity is, thus, proposed. Our model suggests that OTA hepatotoxicity compromises detoxification and antioxidant pathways, leading to mitochondrial membrane dysfunction manifested by crosstalk between pathways of energy metabolism. Interestingly, our data additionally aligns with a possible role of mitochondrial fusion as a "passive mechanism" to rescue mitochondrial integrity during OTA toxicity.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
-
- Boutrif E, Canet C. Mycotoxin prevention and control: FAO programmes. Revue De Medecine Veterinaire. 1998;149:681–694.
-
- Binder E. Managing the risk of mycotoxins in modern feed production. Anim. Feed Sci. Technol. 2007;133:149–166. doi: 10.1016/j.anifeedsci.2006.08.008. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
