

The cytokines interleukin-6 and interferon-α induce distinct microglia phenotypes
- PMID: 35429976
- PMCID: PMC9013466
- DOI: 10.1186/s12974-022-02441-x
The cytokines interleukin-6 and interferon-α induce distinct microglia phenotypes
Abstract
Background: Elevated production of the cytokines interleukin (IL)-6 or interferon (IFN)-α in the central nervous system (CNS) is implicated in the pathogenesis of neurological diseases such as neuromyelitis optica spectrum disorders or cerebral interferonopathies, respectively. Transgenic mice with CNS-targeted chronic production of IL-6 (GFAP-IL6) or IFN-α (GFAP-IFN) recapitulate important clinical and pathological features of these human diseases. The activation of microglia is a prominent manifestation found both in the human diseases and in the transgenic mice, yet little is known about how this contributes to disease pathology.
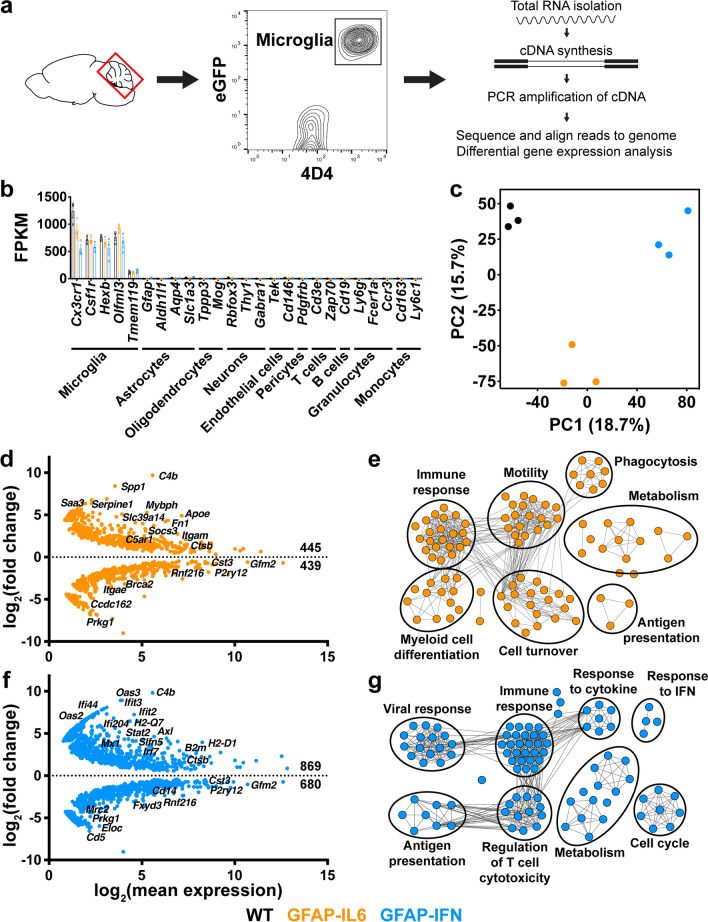
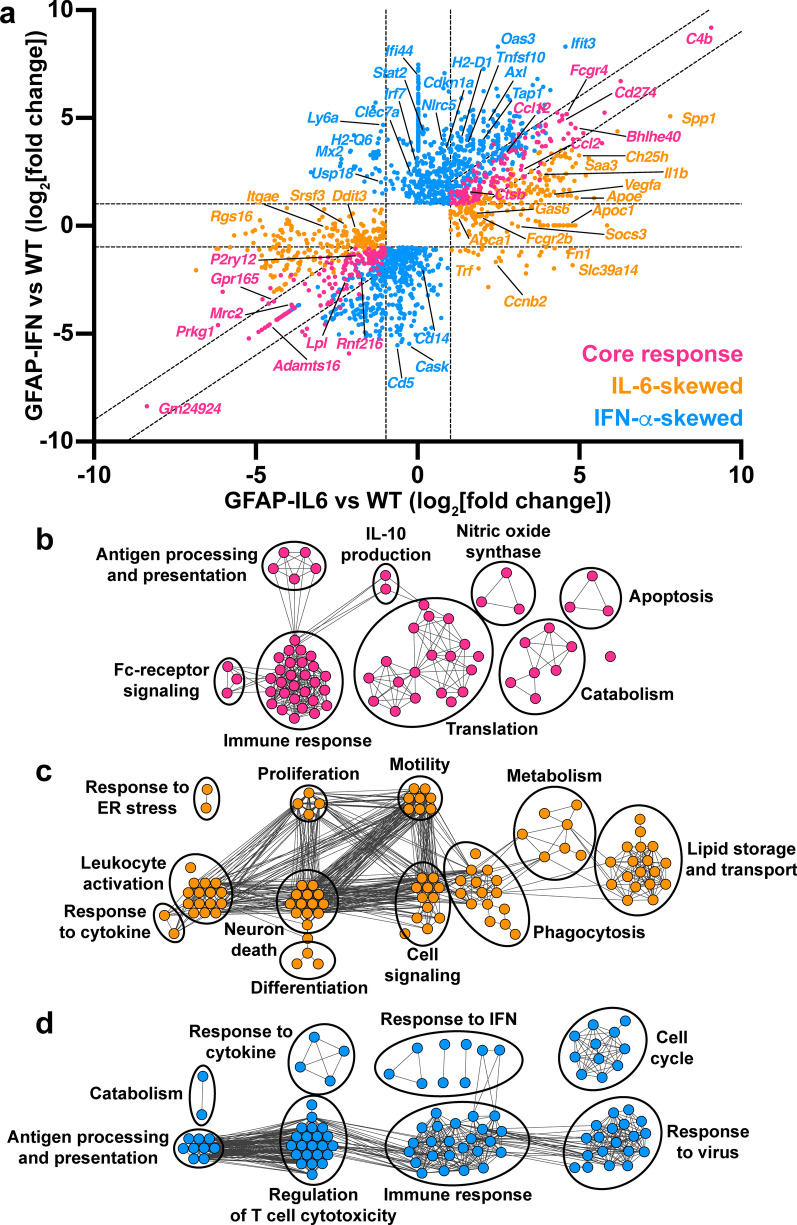
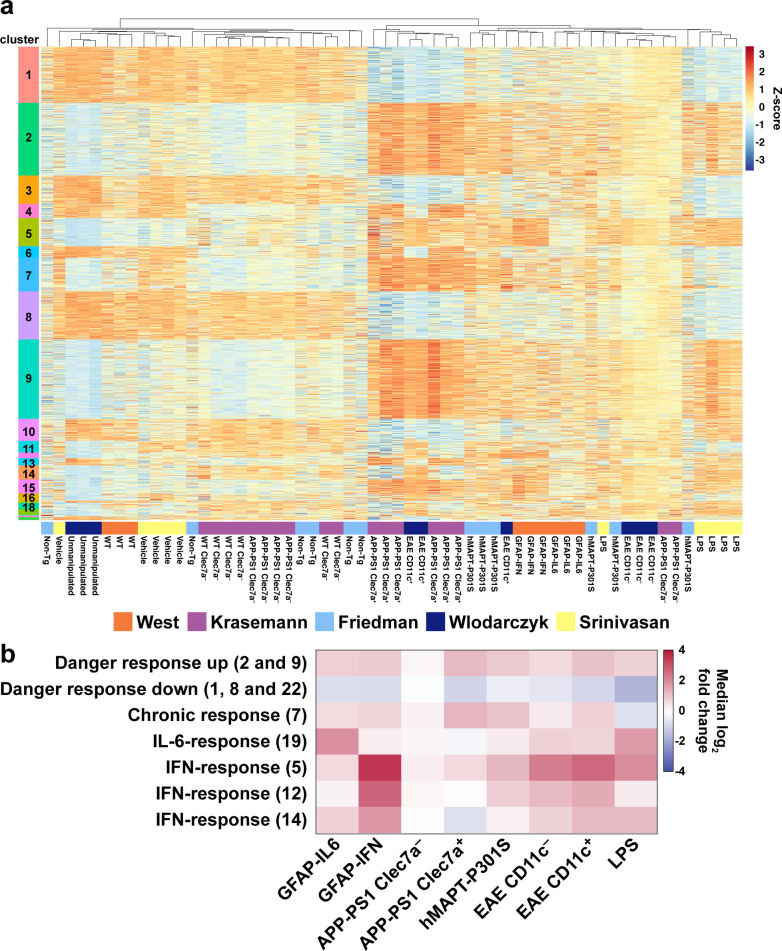
Methods: Here, we used a combination of ex vivo and in situ techniques to characterize the molecular, cellular and transcriptomic phenotypes of microglia in GFAP-IL6 versus GFAP-IFN mice. In addition, a transcriptomic meta-analysis was performed to compare the microglia response from GFAP-IL6 and GFAP-IFN mice to the response of microglia in a range of neurodegenerative and neuroinflammatory disorders.
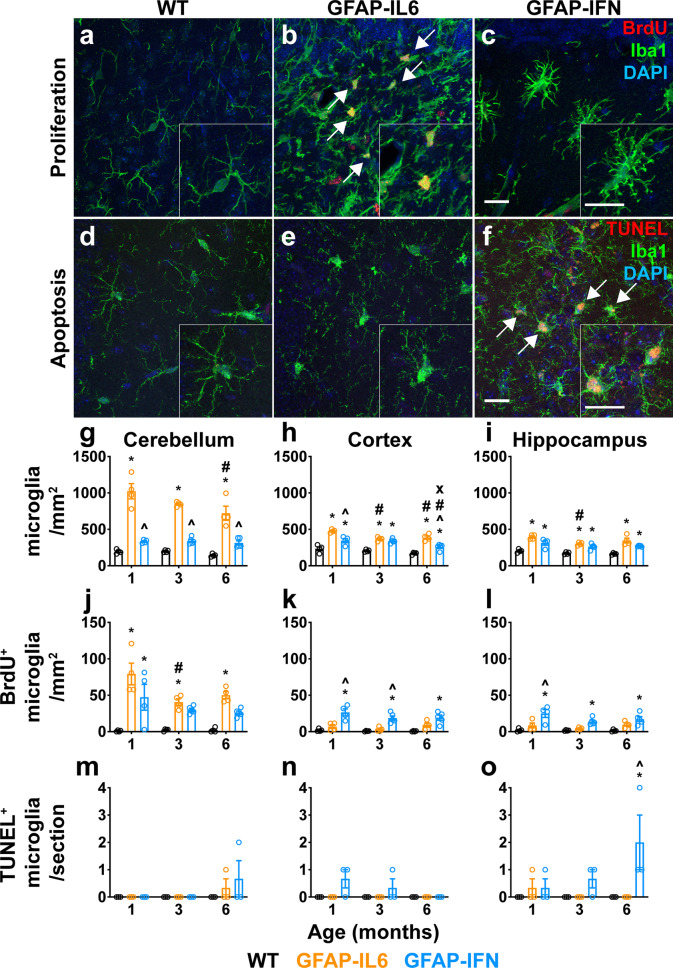
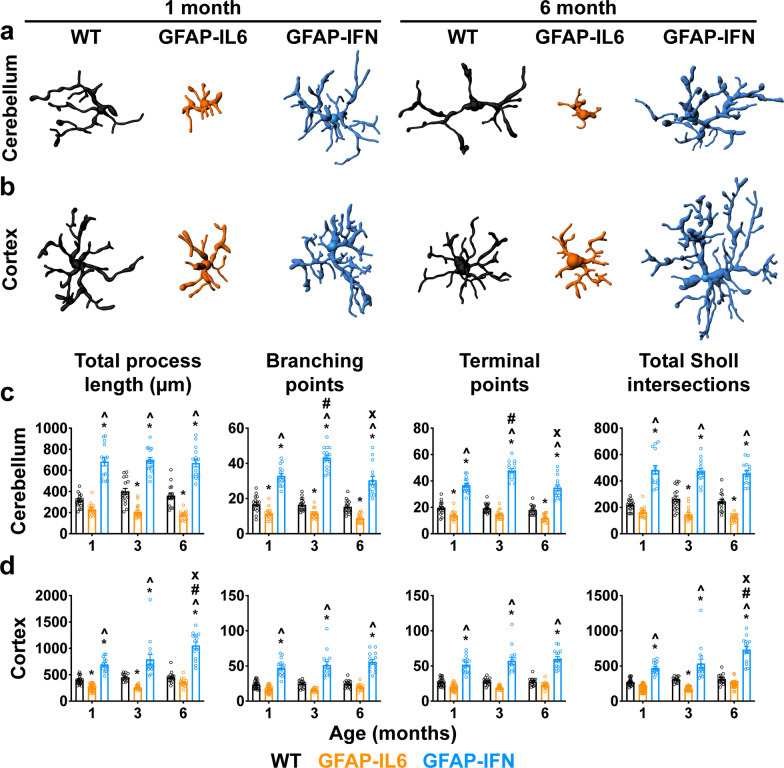
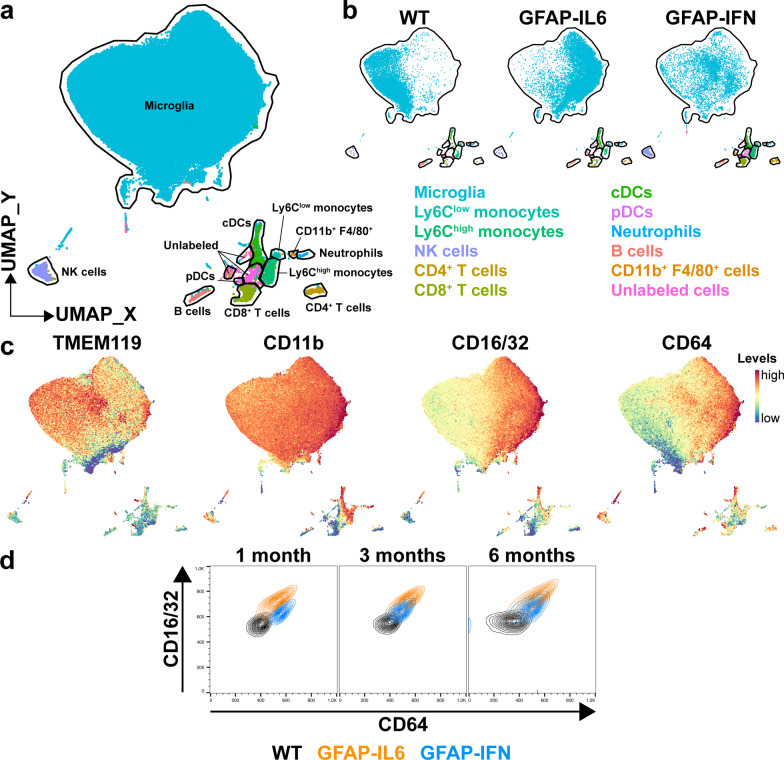
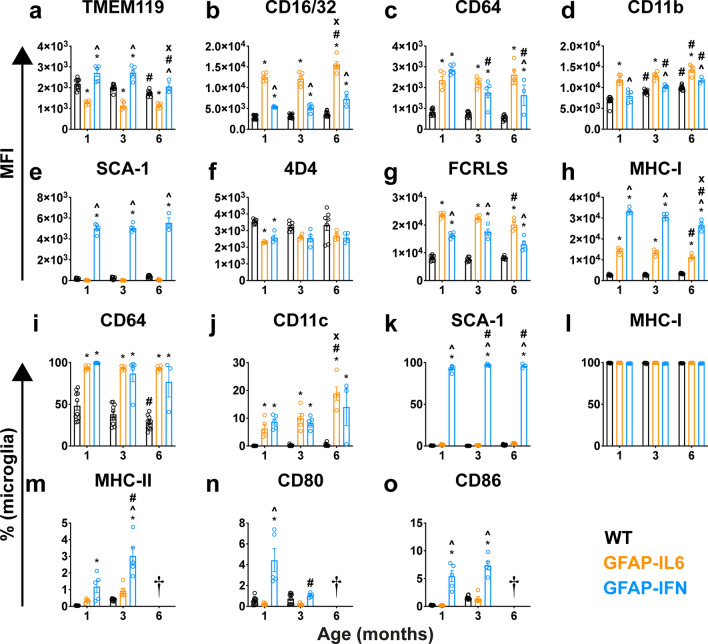
Results: We demonstrated that microglia show stimulus-specific responses to IL-6 versus IFN-α in the brain resulting in unique and extensive molecular and cellular adaptations. In GFAP-IL6 mice, microglia proliferated, had shortened, less branched processes and elicited transcriptomic and molecular changes associated with phagocytosis and lipid processing. In comparison, microglia in the brain of GFAP-IFN mice exhibited increased proliferation and apoptosis, had larger, hyper-ramified processes and showed transcriptomic and surface marker changes associated with antigen presentation and antiviral response. Further, a transcriptomic meta-analysis revealed that IL-6 and IFN-α both contribute to the formation of a core microglia response in animal models of neurodegenerative and neuroinflammatory disorders, such as Alzheimer's disease, tauopathy, multiple sclerosis and lipopolysaccharide-induced endotoxemia.
Conclusions: Our findings demonstrate that microglia responses to IL-6 and IFN-α are highly stimulus-specific, wide-ranging and give rise to divergent phenotypes that modulate microglia responses in neuroinflammatory and neurodegenerative diseases.
Keywords: Central nervous system; Cytokine; Interferon-alpha; Interleukin-6; Microglia; Neuroinflammation; Phenotype.
© 2022. The Author(s).
Conflict of interest statement
B.G. is a director of Pacific Analytics PTY LTD & SMRTR PTY LTD, Australia, which had no role in the design, execution, analysis or preparation of the manuscript. O.B. has collaborations with Sanofi, GSK, Regulus Therapeutics; research funding from Sanofi, GSK, miRagen Therapeutics, honoraria for lectures, consultancy: Camp4. The other authors declare no other competing interests.
Figures
References
-
- West PK, Viengkhou B, Campbell IL, Hofer MJ. Microglia responses to interleukin-6 and type I interferons in neuroinflammatory disease. Glia. 2019;67(10):1821–1841. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
