

Ecological and genomic responses of soil microbiomes to high-severity wildfire: linking community assembly to functional potential
- PMID: 35430593
- PMCID: PMC9213548
- DOI: 10.1038/s41396-022-01232-9
Ecological and genomic responses of soil microbiomes to high-severity wildfire: linking community assembly to functional potential
Abstract
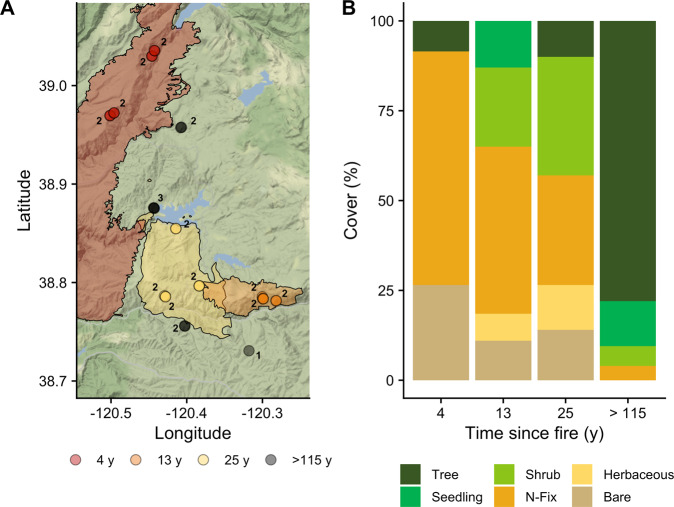
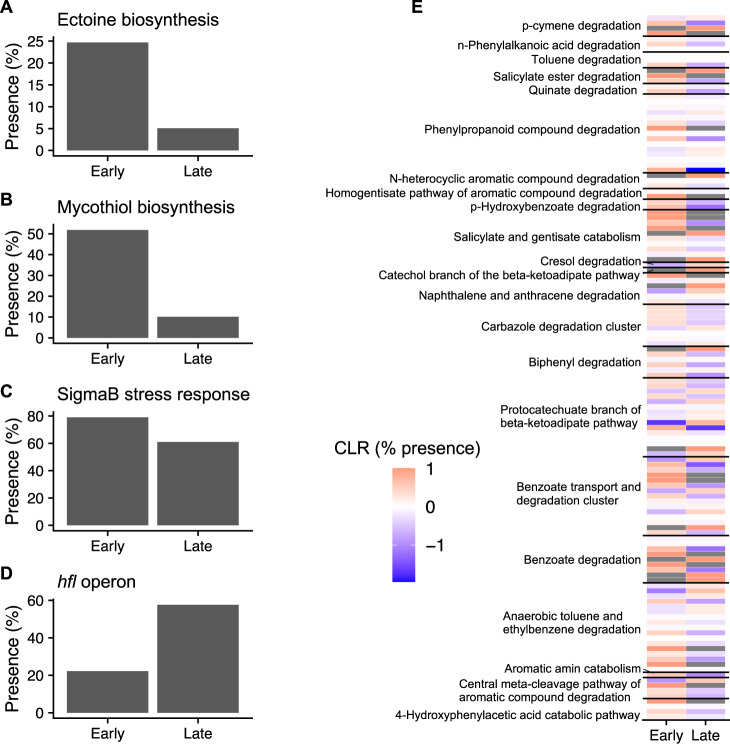
Increasing wildfire severity, which is common throughout the western United States, can have deleterious effects on plant regeneration and large impacts on carbon (C) and nitrogen (N) cycling rates. Soil microbes are pivotal in facilitating these elemental cycles, so understanding the impact of increasing fire severity on soil microbial communities is critical. Here, we assess the long-term impact of high-severity fires on the soil microbiome. We find that high-severity wildfires result in a multi-decadal (>25 y) recovery of the soil microbiome mediated by concomitant differences in aboveground vegetation, soil chemistry, and microbial assembly processes. Our results depict a distinct taxonomic and functional successional pattern of increasing selection in post-fire soil microbial communities. Changes in microbiome composition corresponded with changes in microbial functional potential, specifically altered C metabolism and enhanced N cycling potential, which related to rates of potential decomposition and inorganic N availability, respectively. Based on metagenome-assembled genomes, we show that bacterial genomes enriched in our earliest site (4 y since fire) harbor distinct traits such as a robust stress response and a high potential to degrade pyrogenic, polyaromatic C that allow them to thrive in post-fire environments. Taken together, these results provide a biological basis for previously reported process rate measurements and explain the temporal dynamics of post-fire biogeochemistry, which ultimately constrains ecosystem recovery.
© 2022. This is a U.S. government work and not under copyright protection in the U.S.; foreign copyright protection may apply.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
-
- Stephens SL, Burrows N, Buyantuyev A, Gray RW, Keane RE, Kubian R, et al. Temperate and boreal forest mega-fires: characteristics and challenges. Front Ecol Environ. 2014;12:115–22. doi: 10.1890/120332. - DOI
-
- Adams MA. Mega-fires, tipping points and ecosystem services: managing forests and woodlands in an uncertain future. Ecol Manag. 2013;294:250–61. doi: 10.1016/j.foreco.2012.11.039. - DOI
-
- McLauchlan KK, Higuera PE, Miesel J, Rogers BM, Schweitzer J, Shuman JK, et al. Fire as a fundamental ecological process: research advances and frontiers. J Ecol. 2020;108:2047–69. doi: 10.1111/1365-2745.13403. - DOI
-
- Whitman T, Whitman E, Woolet J, Flannigan MD, Thompson DK, Parisien M-A. Soil bacterial and fungal response to wildfires in the Canadian boreal forest across a burn severity gradient. Soil Biol Biochem. 2019;138:107571. doi: 10.1016/j.soilbio.2019.107571. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
