

Selenium Supplementation Improved Cardiac Functions by Suppressing DNMT2-Mediated GPX1 Promoter DNA Methylation in AGE-Induced Heart Failure
- PMID: 35432721
- PMCID: PMC9007686
- DOI: 10.1155/2022/5402997
Selenium Supplementation Improved Cardiac Functions by Suppressing DNMT2-Mediated GPX1 Promoter DNA Methylation in AGE-Induced Heart Failure
Abstract
Objective: Advanced glycation end products (AGEs) are featured metabolites associated with diabetic cardiomyopathy which is characterized by heart failure caused by myocyte apoptosis. Selenium was proved cardioprotective. This study was aimed at investigating the therapeutic effects and underlying mechanisms of selenium supplementation on AGE-induced heart failure.
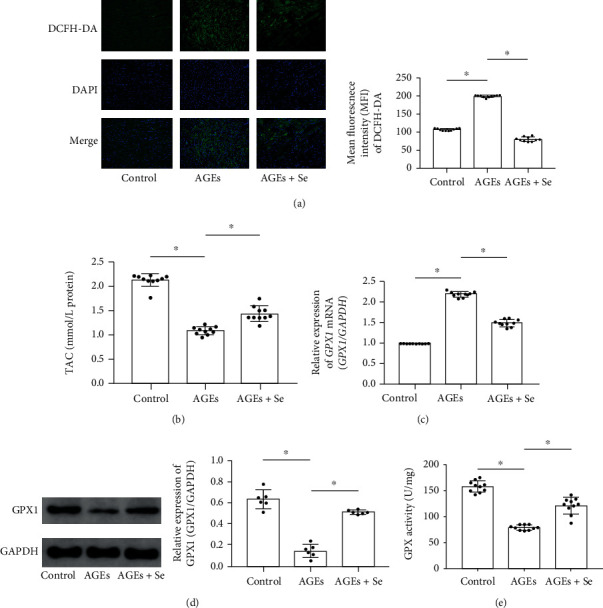
Methods: Rats and primary myocytes were exposed to AGEs. Selenium supplementation was administrated. Cardiac functions and myocyte apoptosis were evaluated. Oxidative stress was assessed by total antioxidant capacity (TAC), reactive oxygen species (ROS) generation, and GPX activity. Expression levels of DNA methyltransferases (DNMTs) and glutathione peroxidase 1 (GPX1) were evaluated. DNA methylation of the GPX1 promoter was analyzed.
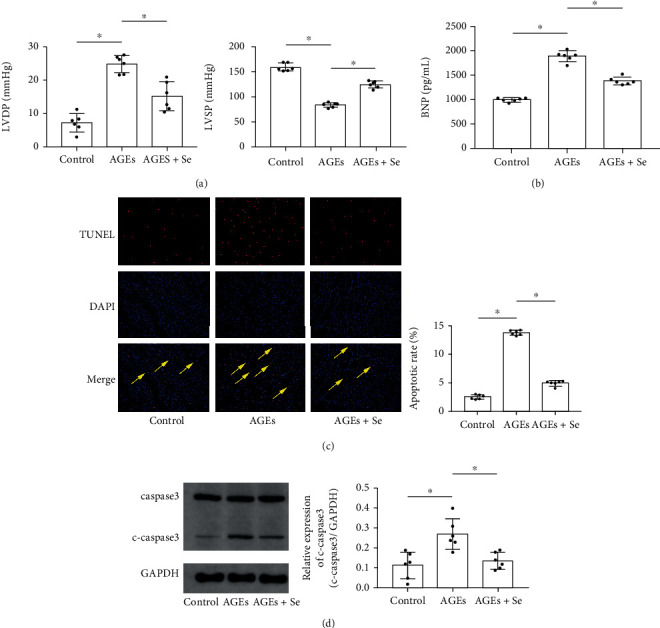
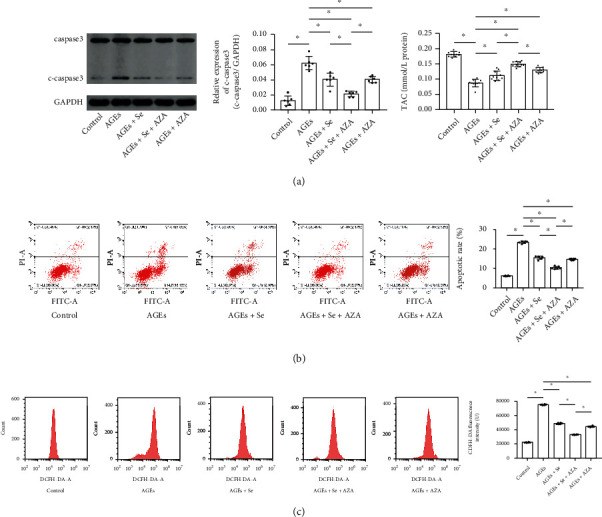
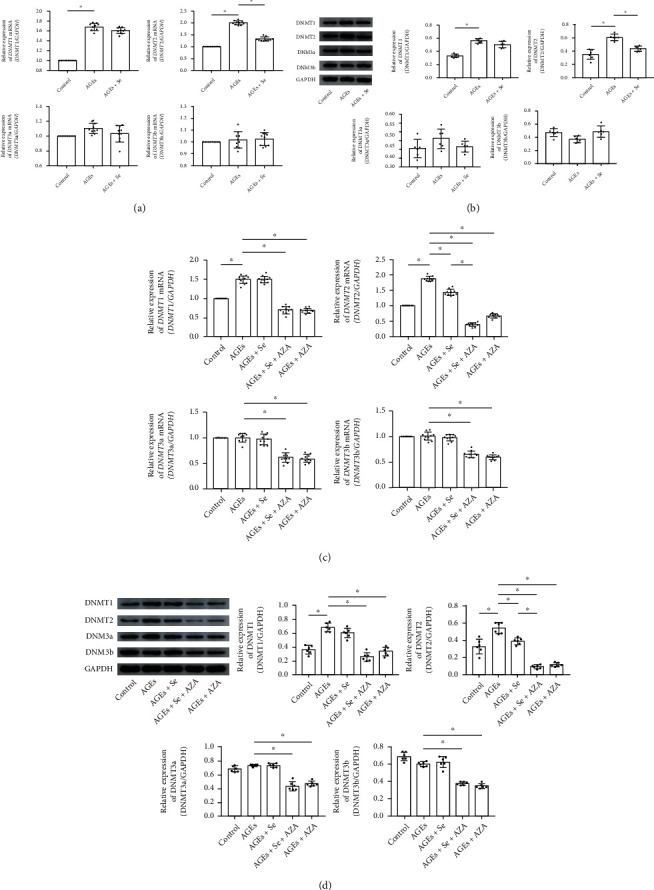
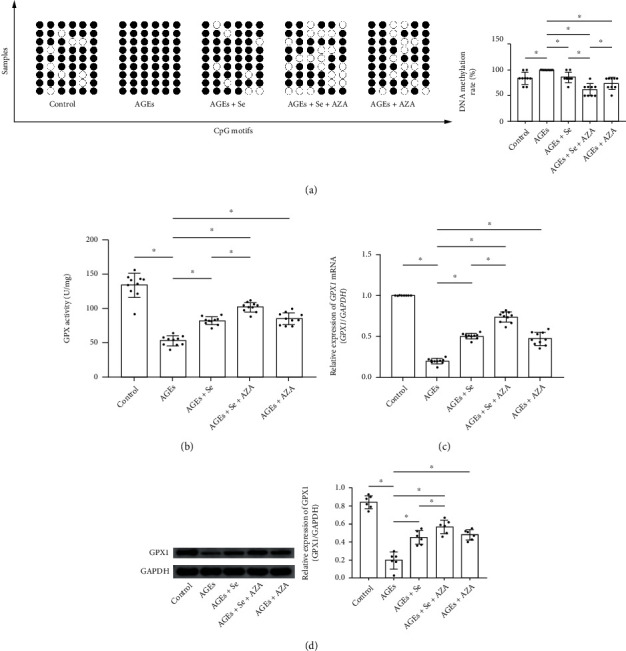
Results: AGE exposure elevated intracellular ROS generation, induced myocyte apoptosis, and impaired cardiac functions. AGE exposure increased DNMT1 and DNMT2 expression, leading to the reduction of GPX1 expression and activity in the heart. Selenium supplementation decreased DNMT2 expression, recovered GPX1 expression and activity, and alleviated intracellular ROS generation and myocyte apoptosis, resulting in cardiac function recovery. DNA methylation analysis in primary myocytes indicated that selenium supplementation or DNMT inhibitor AZA treatment reduced DNA methylation of the GPX1 gene promoter. Selenium supplementation and AZA administration showed synergic inhibitory effect on GPX1 gene promoter methylation.
Conclusions: Selenium supplementation showed cardioprotective effects on AGE-induced heart failure by suppressing ROS-mediated myocyte apoptosis. Selenium supplementation suppressed ROS generation by increasing GPX1 expression via inhibiting DNMT2-induced GPX1 gene promoter DNA methylation in myocytes exposed to AGEs.
Copyright © 2022 Huolan Zhu et al.
Conflict of interest statement
All authors declare no conflicts of interest.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
