

Cell-Selective Adeno-Associated Virus-Mediated SCN1A Gene Regulation Therapy Rescues Mortality and Seizure Phenotypes in a Dravet Syndrome Mouse Model and Is Well Tolerated in Nonhuman Primates
- PMID: 35435735
- PMCID: PMC9242722
- DOI: 10.1089/hum.2022.037
Cell-Selective Adeno-Associated Virus-Mediated SCN1A Gene Regulation Therapy Rescues Mortality and Seizure Phenotypes in a Dravet Syndrome Mouse Model and Is Well Tolerated in Nonhuman Primates
Abstract
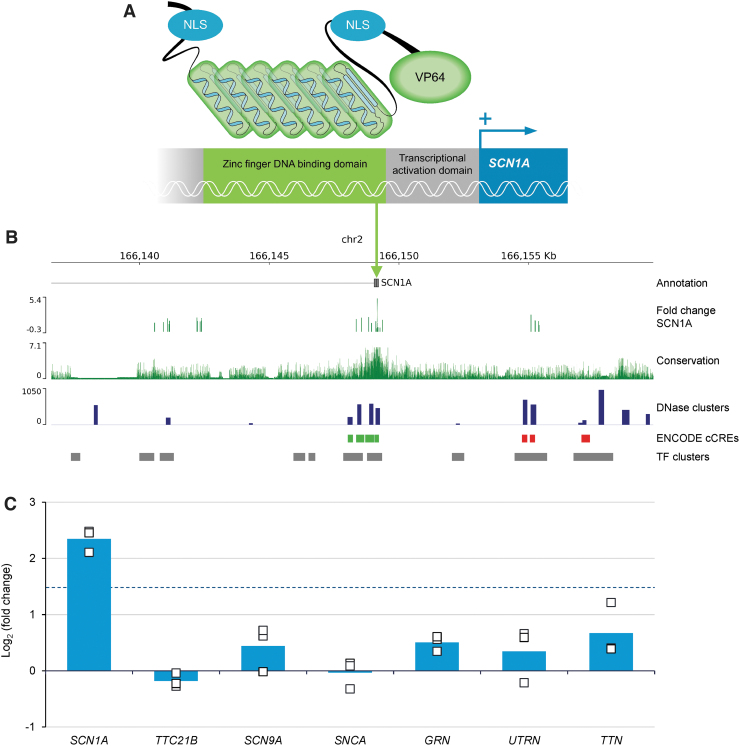
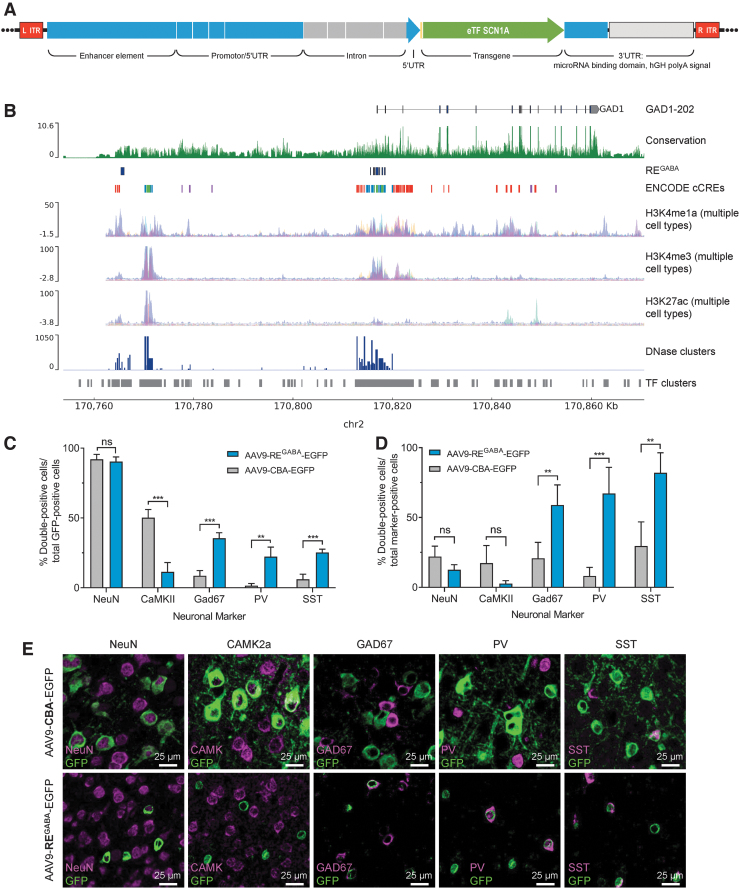
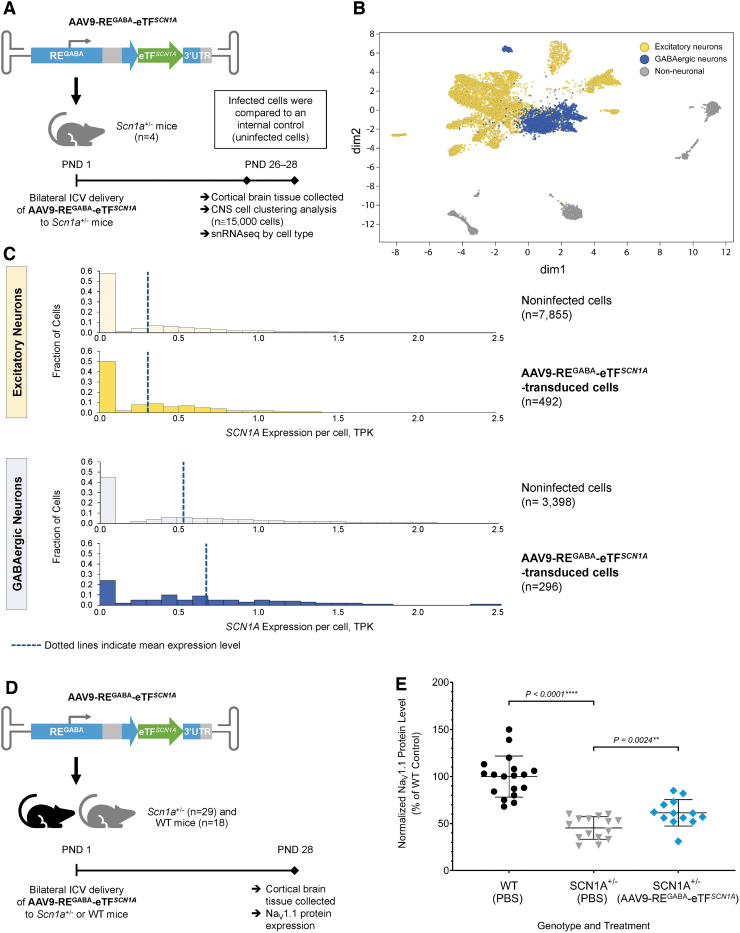
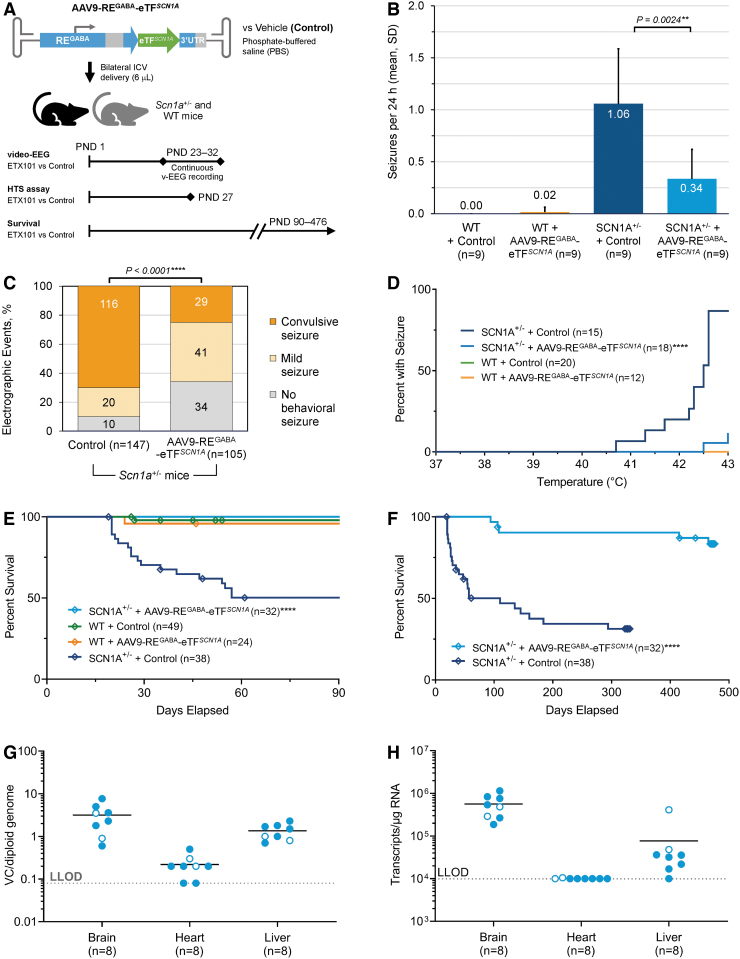
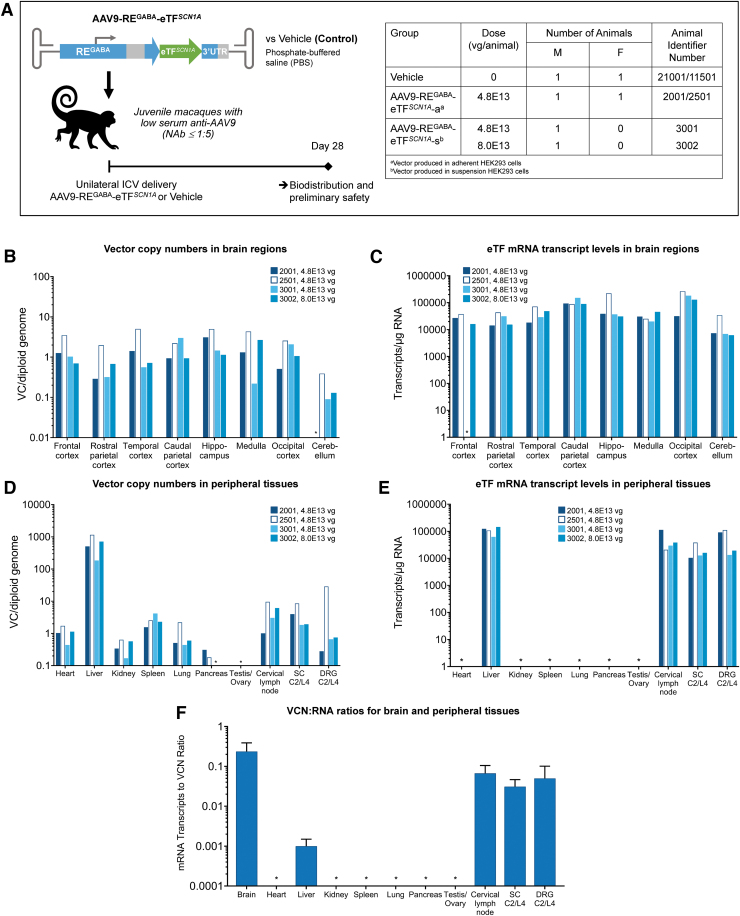
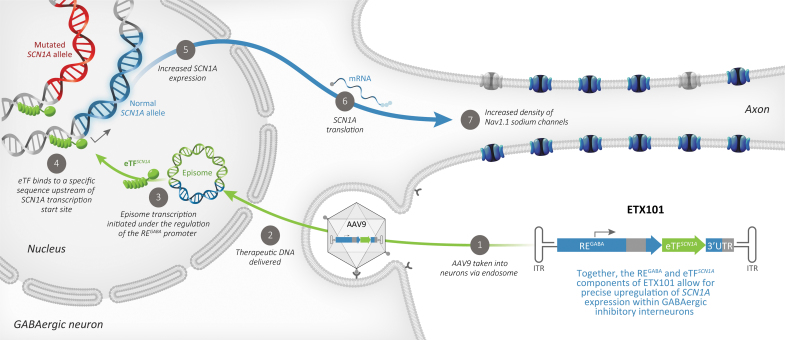
Dravet syndrome (DS) is a developmental and epileptic encephalopathy caused by monoallelic loss-of-function variants in the SCN1A gene. SCN1A encodes for the alpha subunit of the voltage-gated type I sodium channel (NaV1.1), the primary voltage-gated sodium channel responsible for generation of action potentials in GABAergic inhibitory interneurons. In these studies, we tested the efficacy of an adeno-associated virus serotype 9 (AAV9) SCN1A gene regulation therapy, AAV9-REGABA-eTFSCN1A, designed to target transgene expression to GABAergic inhibitory neurons and reduce off-target expression within excitatory cells, in the Scn1a+/- mouse model of DS. Biodistribution and preliminary safety were evaluated in nonhuman primates (NHPs). AAV9-REGABA-eTFSCN1A was engineered to upregulate SCN1A expression levels within GABAergic inhibitory interneurons to correct the underlying haploinsufficiency and circuit dysfunction. A single bilateral intracerebroventricular (ICV) injection of AAV9-REGABA-eTFSCN1A in Scn1a+/- postnatal day 1 mice led to increased SCN1A mRNA transcripts, specifically within GABAergic inhibitory interneurons, and NaV1.1 protein levels in the brain. This was associated with a significant decrease in the occurrence of spontaneous and hyperthermia-induced seizures, and prolonged survival for over a year. In NHPs, delivery of AAV9-REGABA-eTFSCN1A by unilateral ICV injection led to widespread vector biodistribution and transgene expression throughout the brain, including key structures involved in epilepsy and cognitive behaviors, such as hippocampus and cortex. AAV9-REGABA-eTFSCN1A was well tolerated, with no adverse events during administration, no detectable changes in clinical observations, no adverse findings in histopathology, and no dorsal root ganglion-related toxicity. Our results support the clinical development of AAV9-REGABA-eTFSCN1A (ETX101) as an effective and targeted disease-modifying approach to SCN1A+ DS.
Keywords: Dravet syndrome; SCN1A; channelopathy; encephalopathy; gene regulation therapy; preclinical models.
Conflict of interest statement
A.T., J.M., R.A., I.W.L., R.H., M.C., J. Li, S.P., M.C.V., M.M., A.B., and S.T. are current employees of and hold stock options in Encoded Therapeutics. A.Y., J.L., T.S., and T.C. are former employees of Encoded Therapeutics. J.A.K. serves as a consultant to Encoded Therapeutics, Praxis Precision Medicines, and NeuroCycle Therapeutics, and serves on the Scientific Advisory Boards of the Dravet Syndrome Foundation and FamilieSCN2A Foundation.
Figures
References
-
- Dravet C, Guerrini R. Dravet Syndrome. Topics in Epilepsy, 3rd ed. Paris: John Libbey Eurotext, 2011.
-
- Brunklaus A, Ellis R, Reavey E, et al. . Prognostic, clinical and demographic features in SCN1A mutation-positive Dravet syndrome. Brain 2012;135(Pt 8):2329–2336. - PubMed
-
- Gataullina S, Dulac O. From genotype to phenotype in Dravet disease. Seizure 2017;44:58–64. - PubMed
-
- Wirrell EC, Laux L, Donner E, et al. . Optimizing the diagnosis and management of Dravet syndrome: recommendations from a North American Consensus Panel. Pediatr Neurol 2017;68:18–34.e3. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Other Literature Sources