

Phosphate dysregulation via the XPR1-KIDINS220 protein complex is a therapeutic vulnerability in ovarian cancer
- PMID: 35437317
- PMCID: PMC9246846
- DOI: 10.1038/s43018-022-00360-7
Phosphate dysregulation via the XPR1-KIDINS220 protein complex is a therapeutic vulnerability in ovarian cancer
Abstract
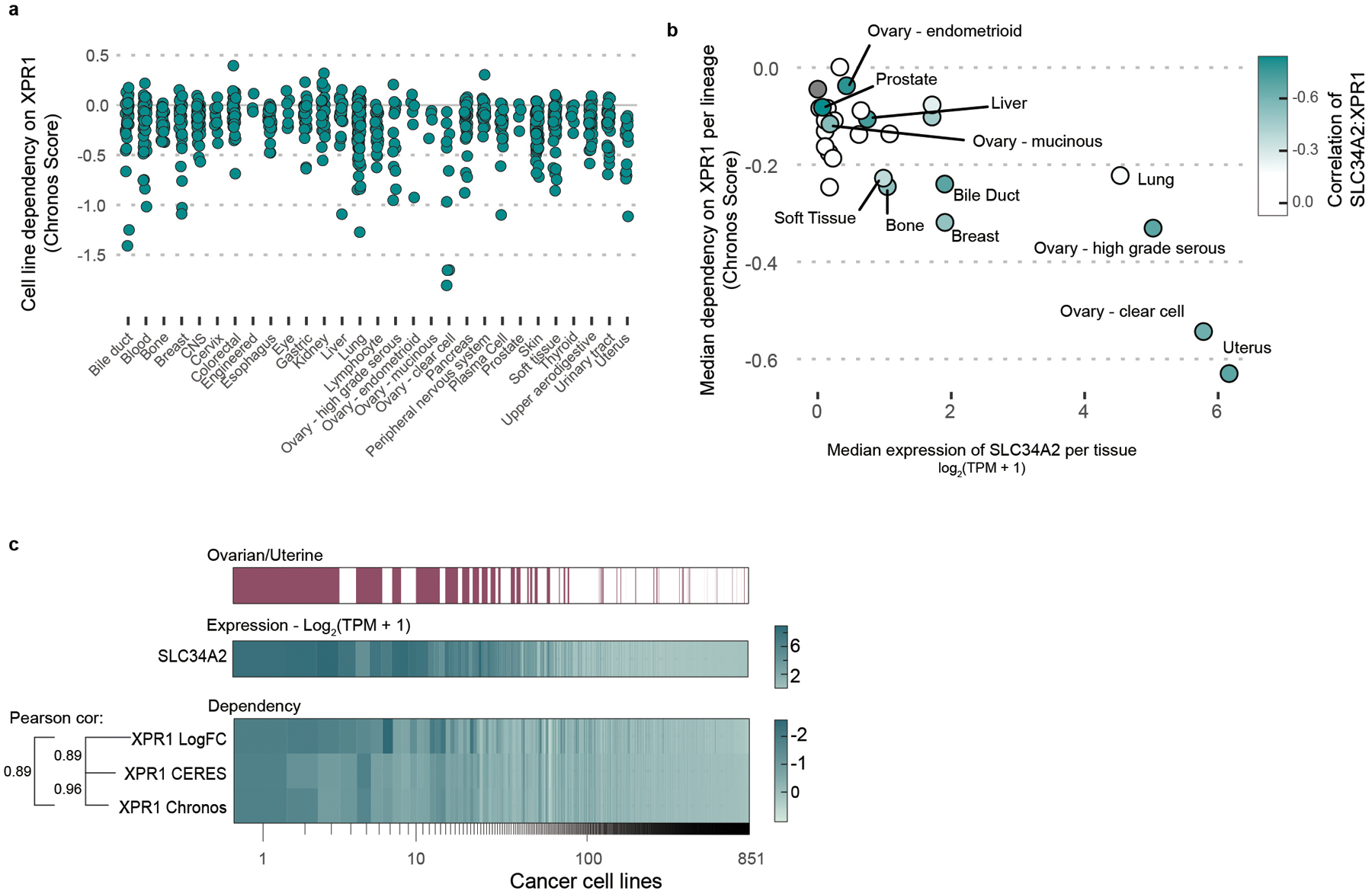
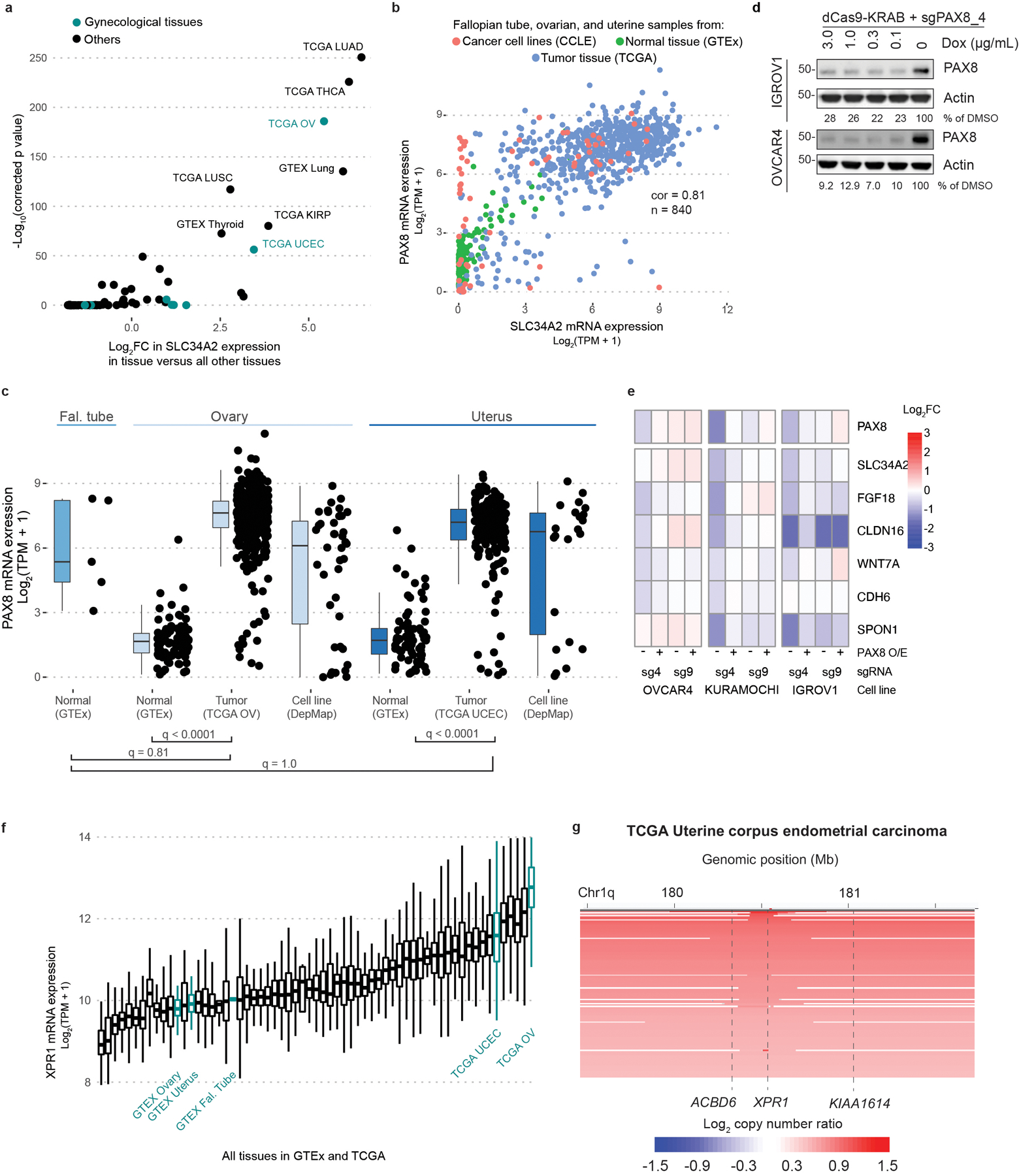
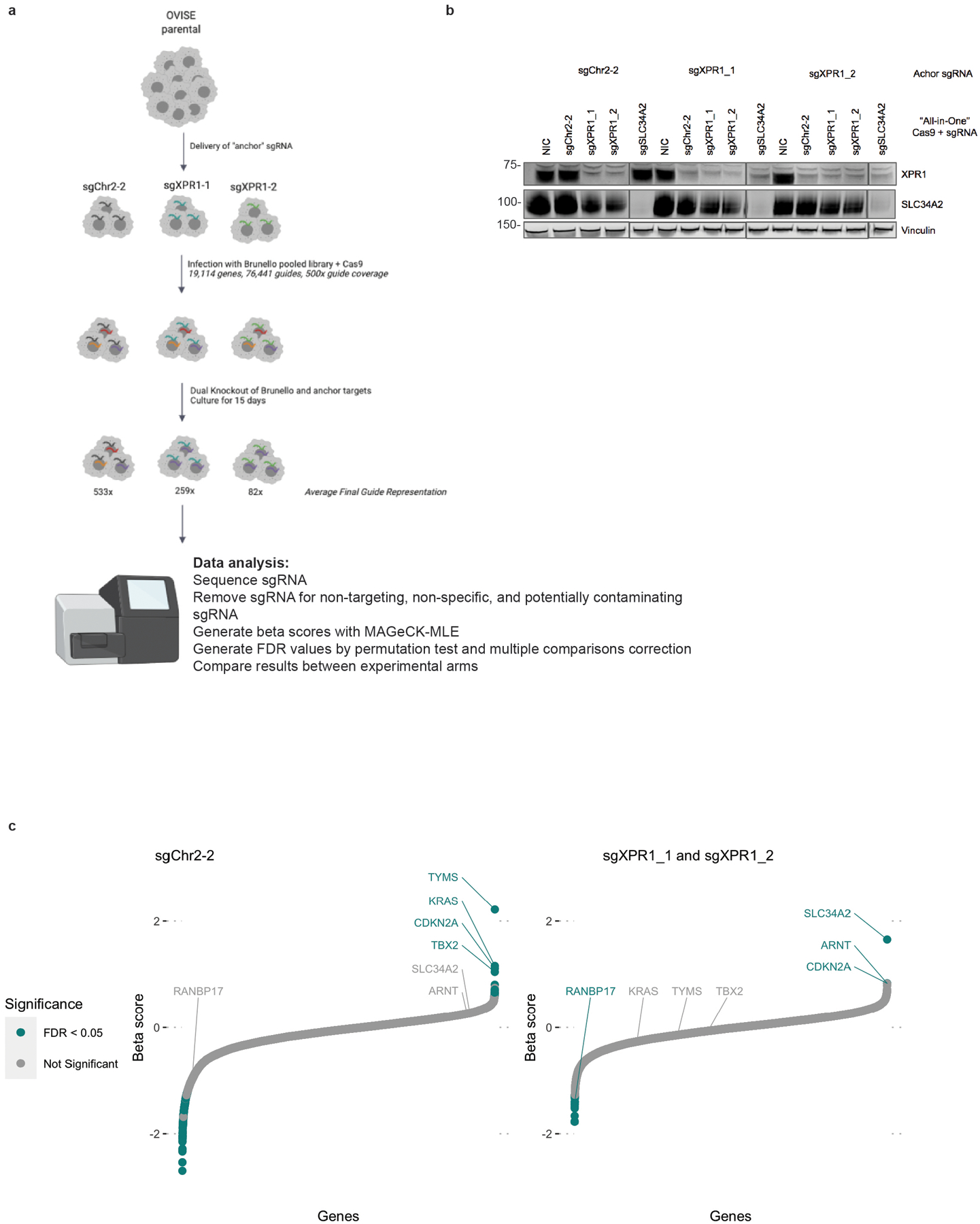
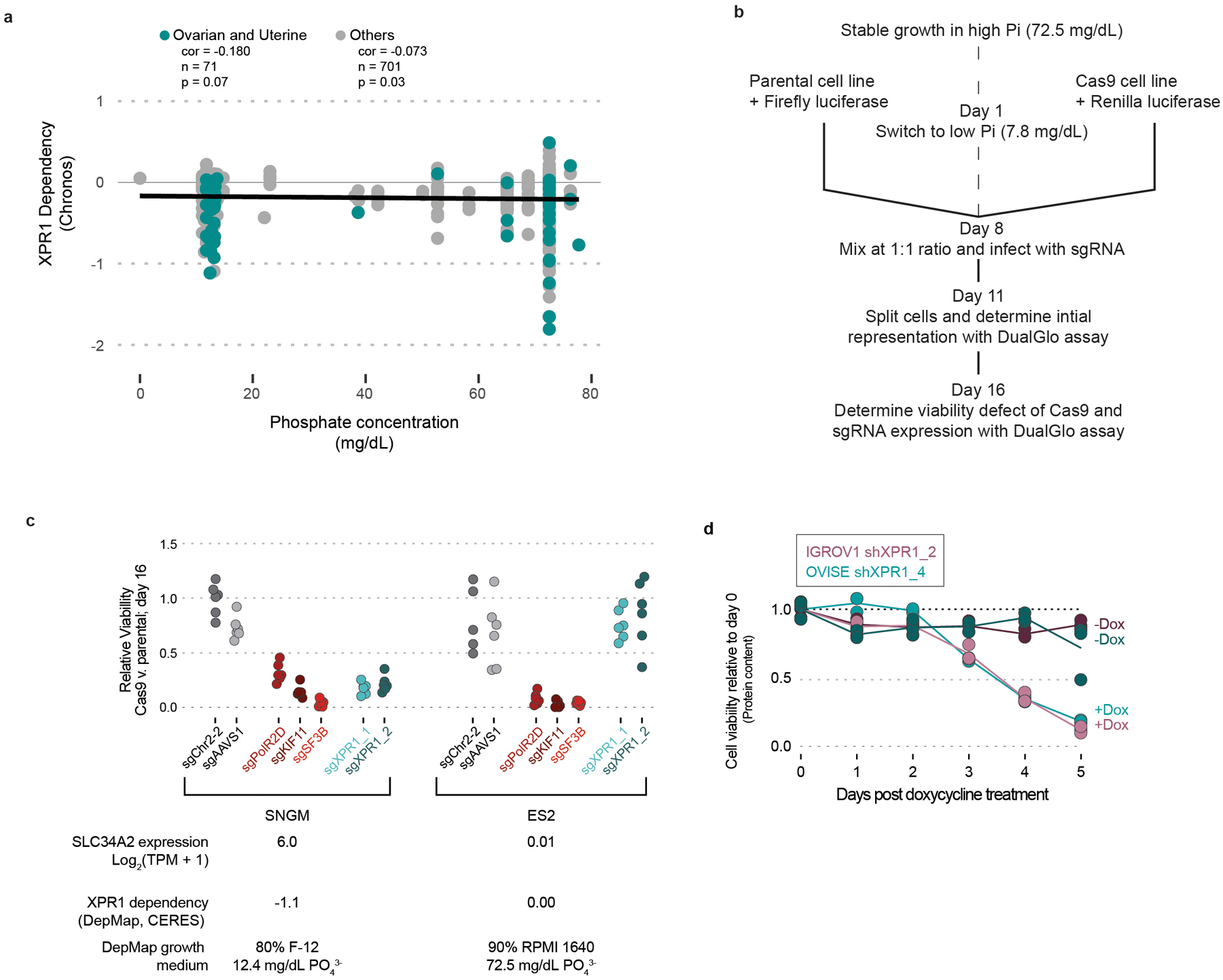
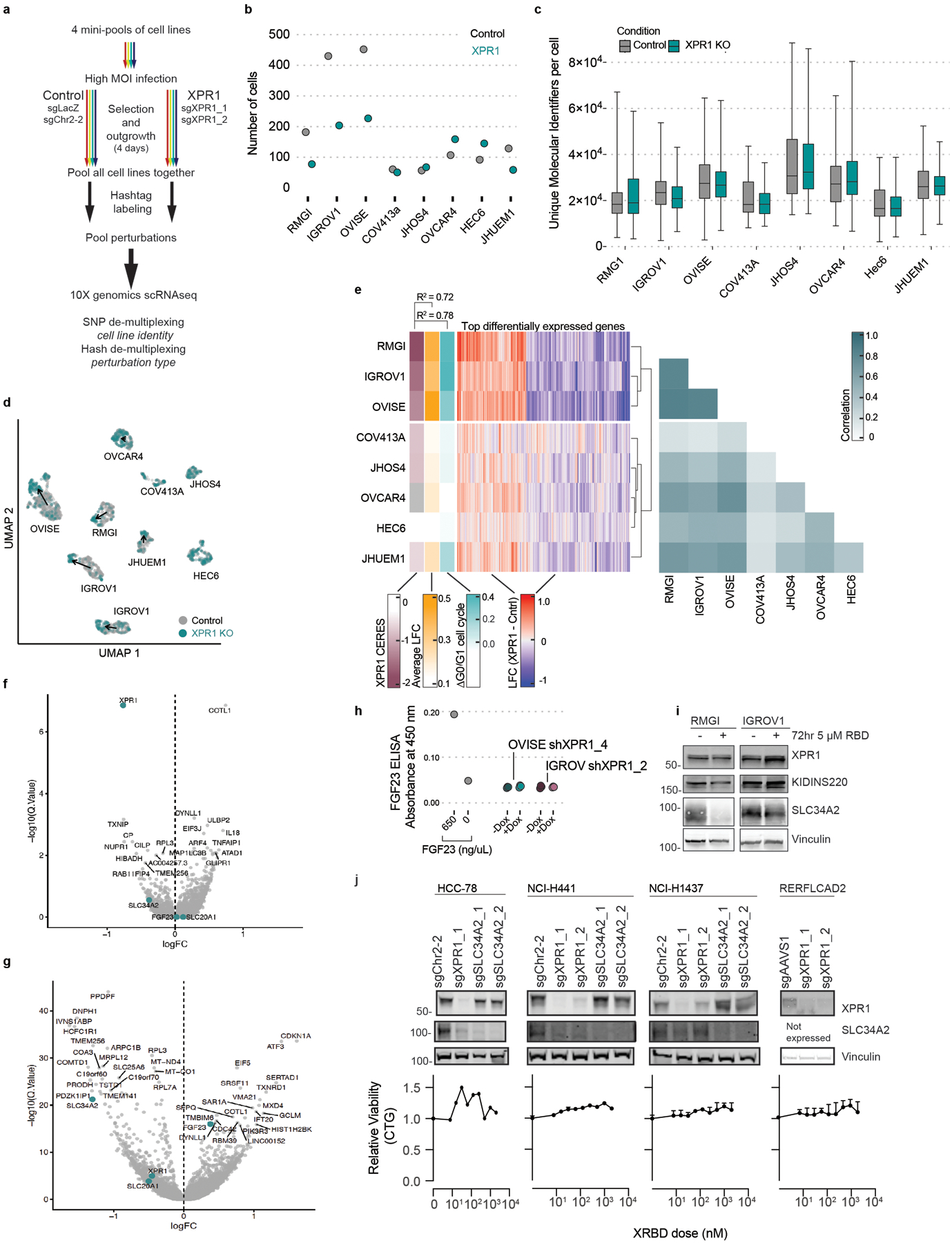
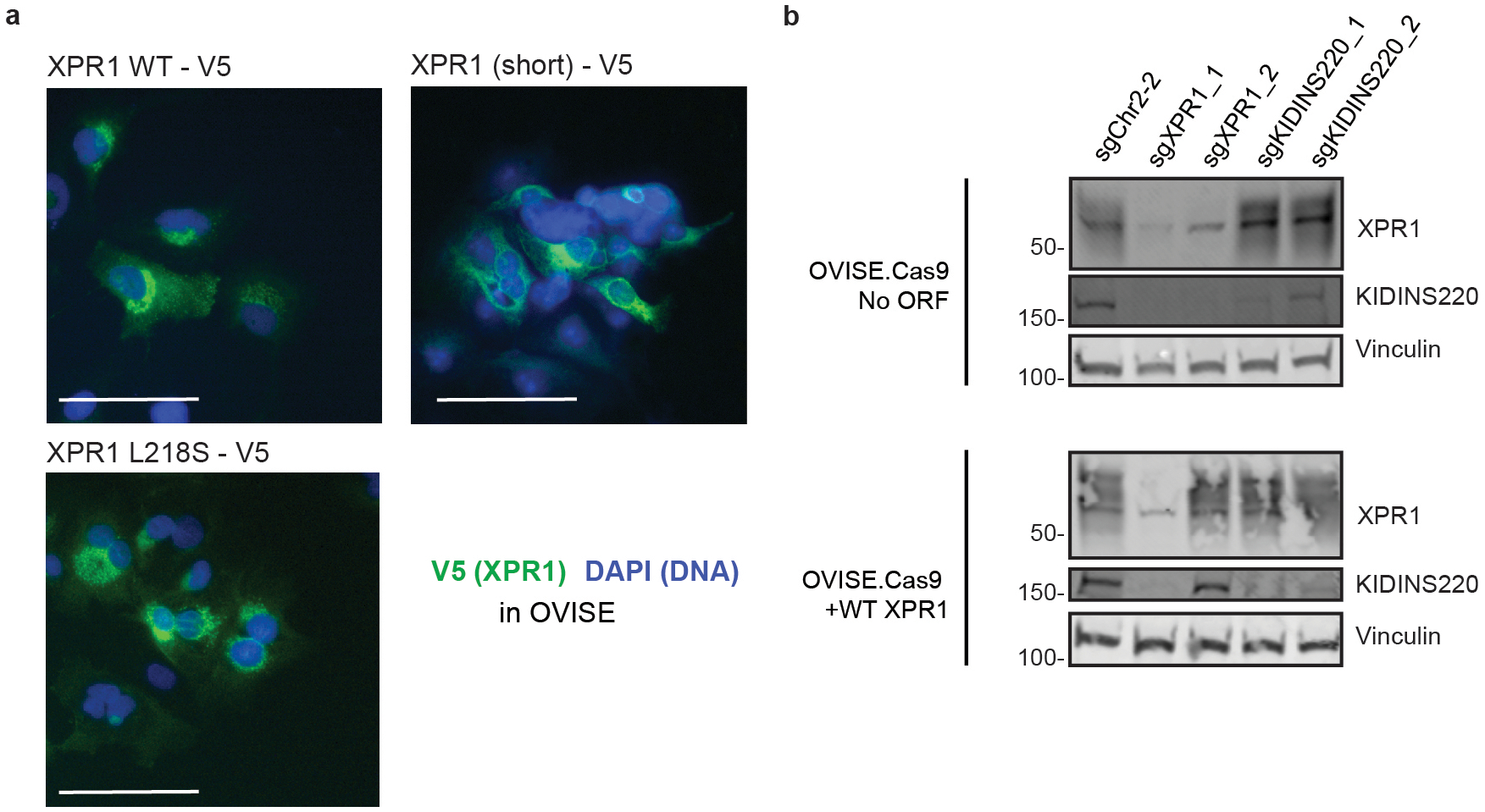
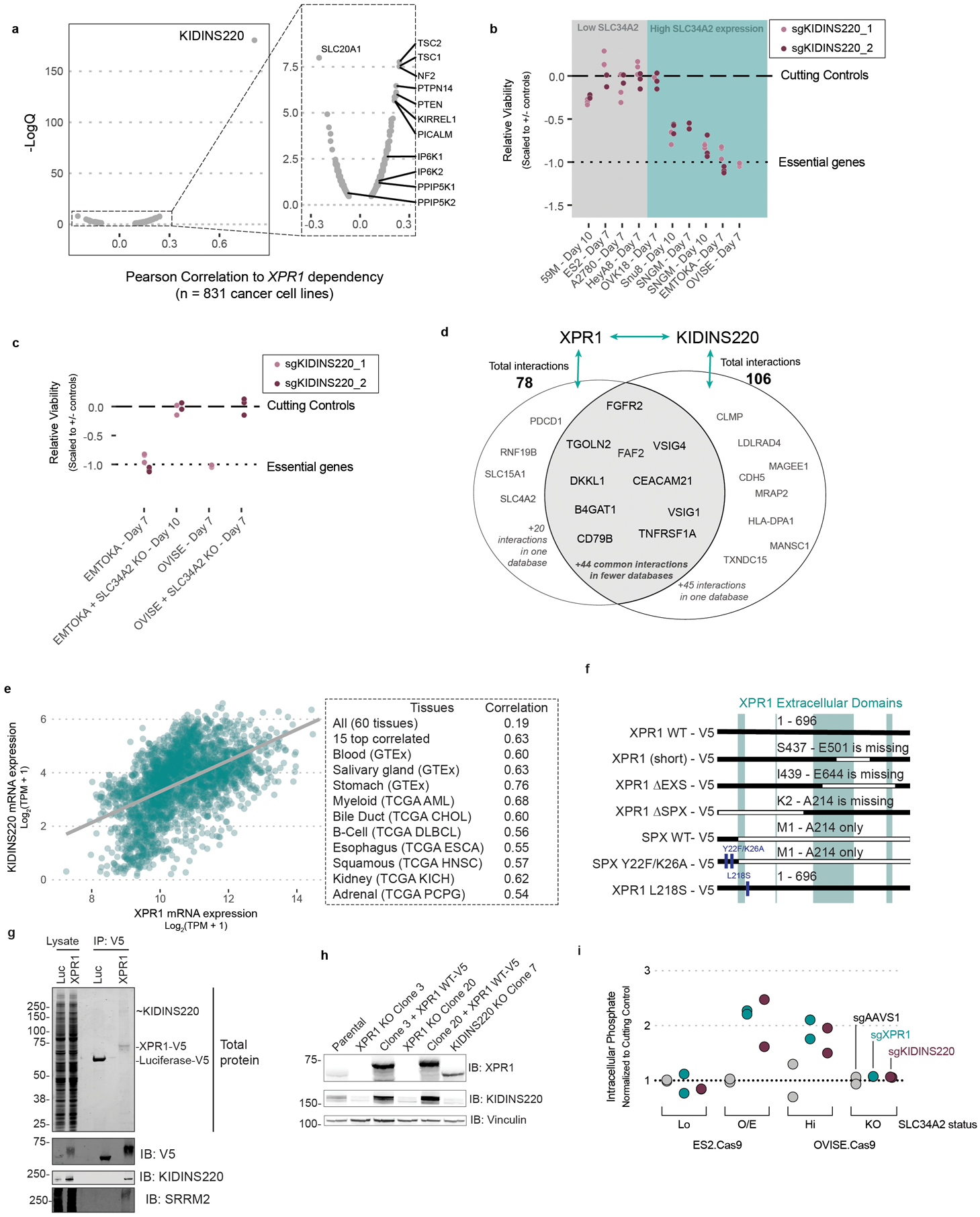
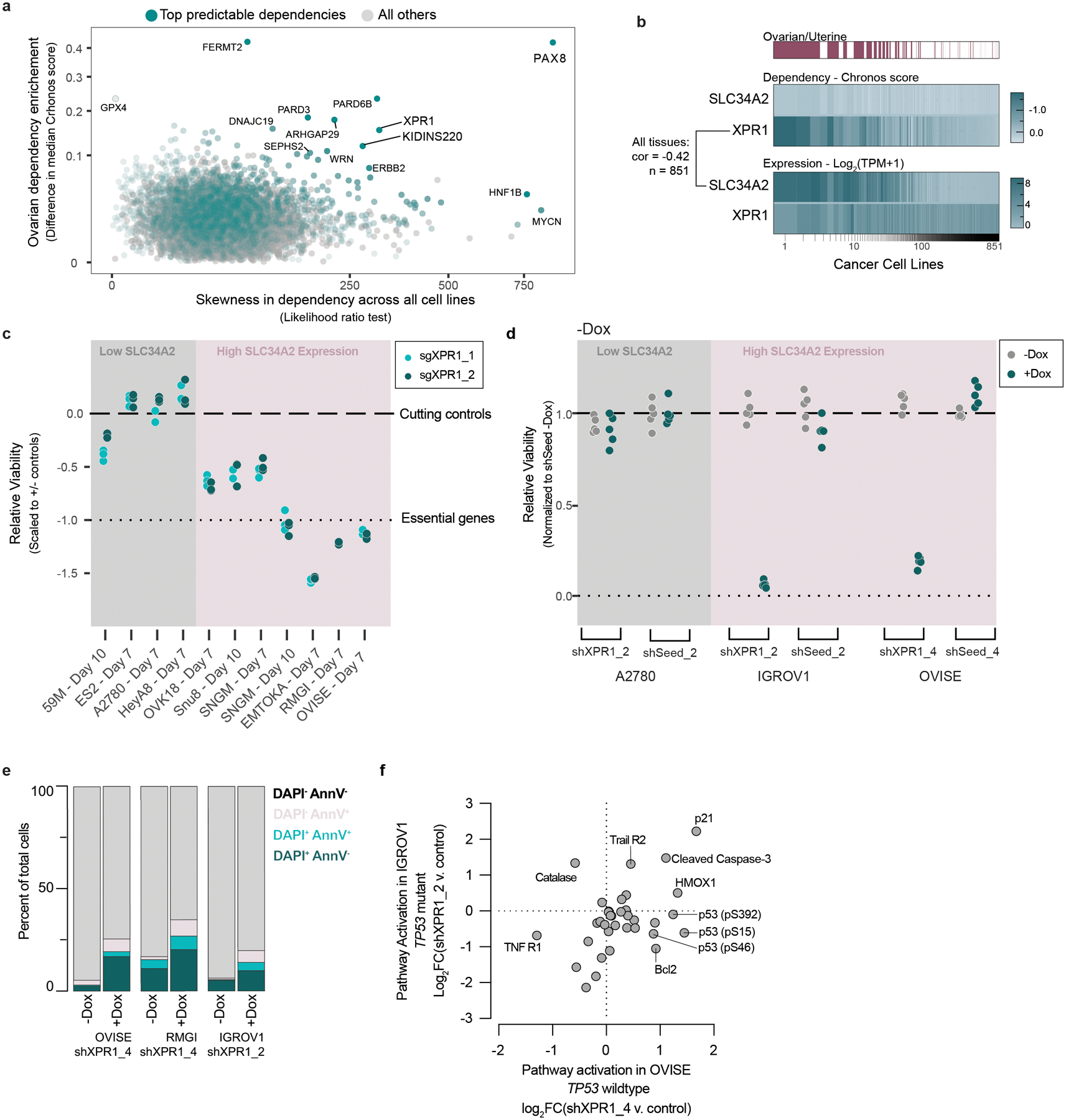
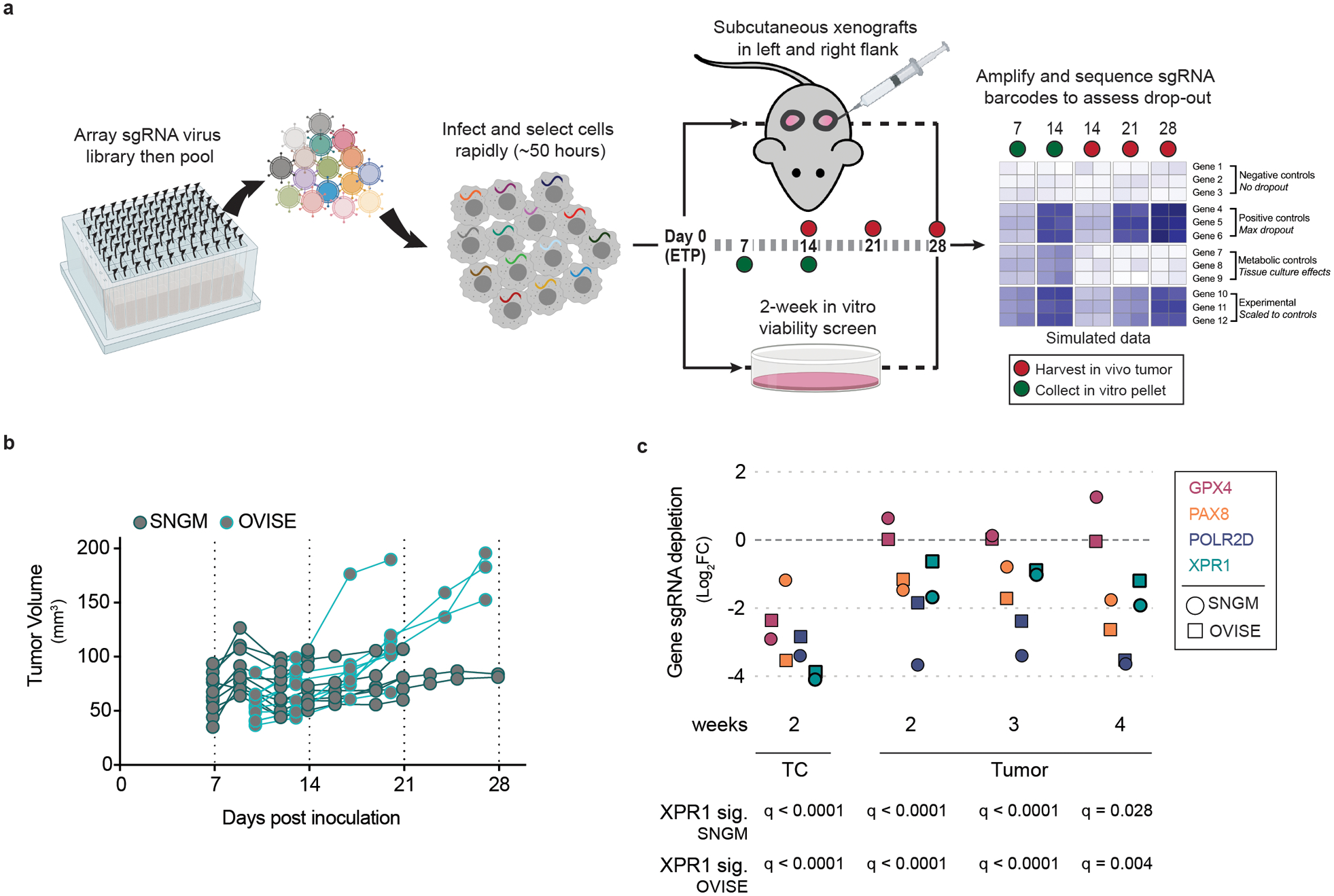
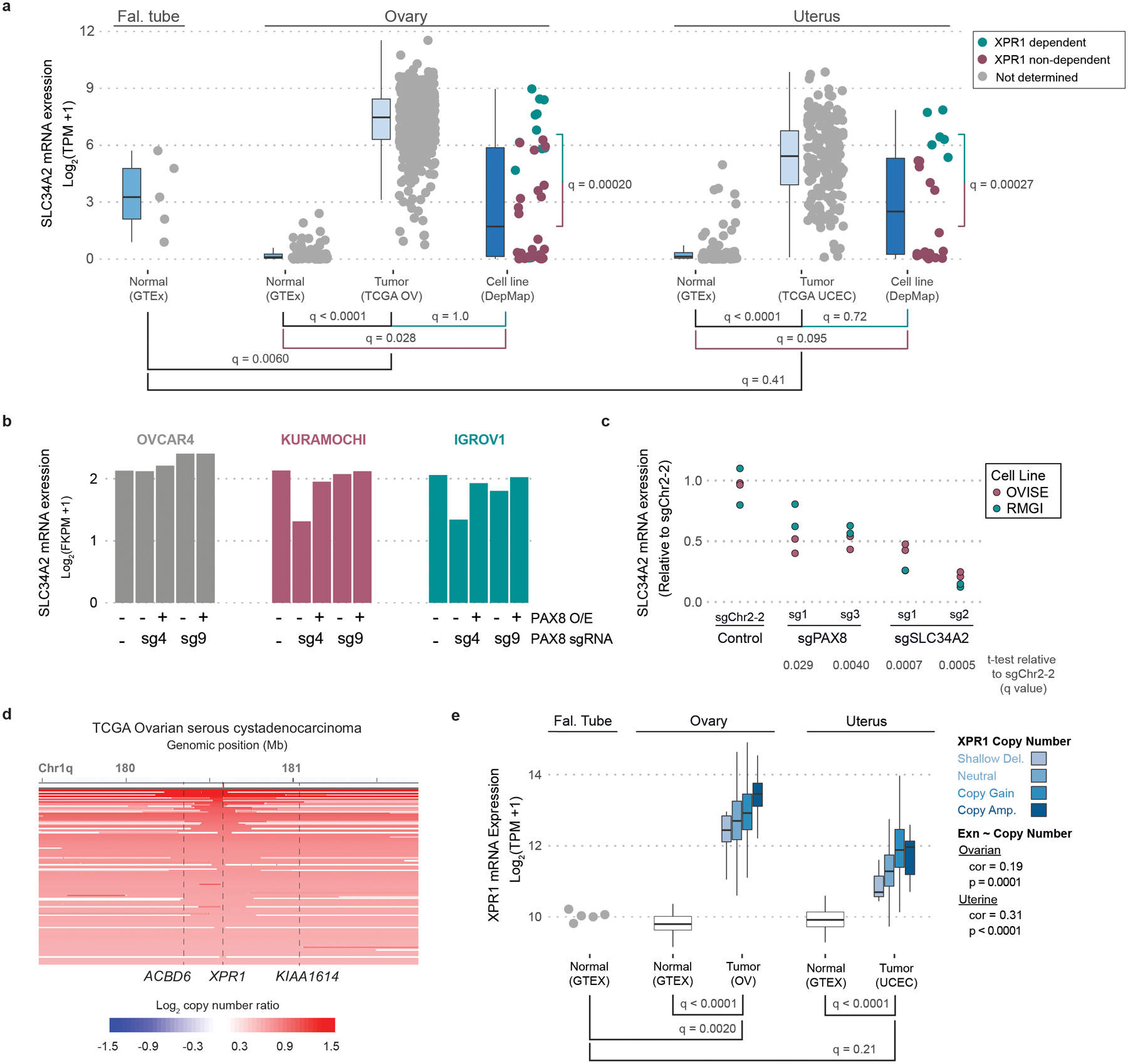
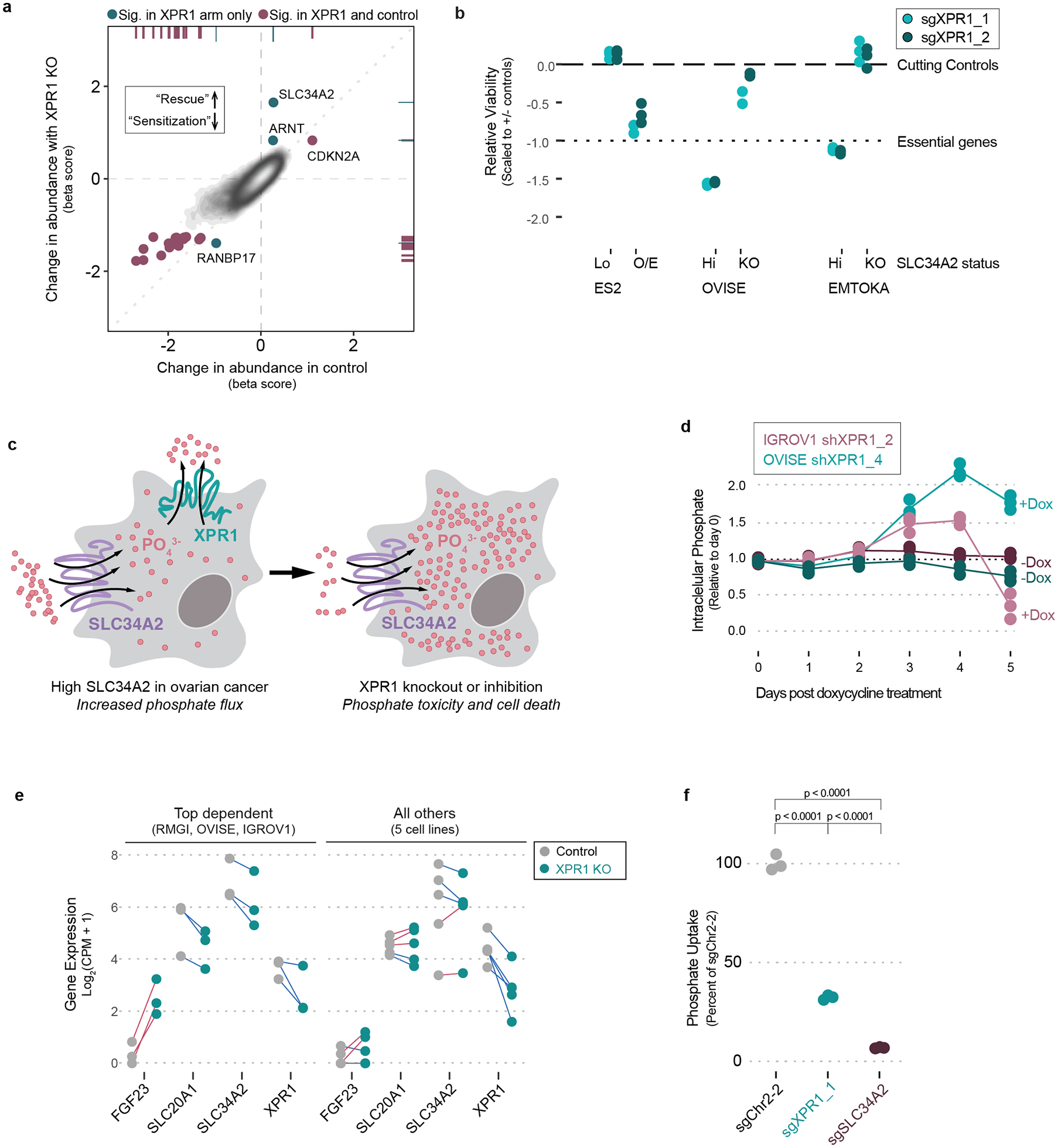
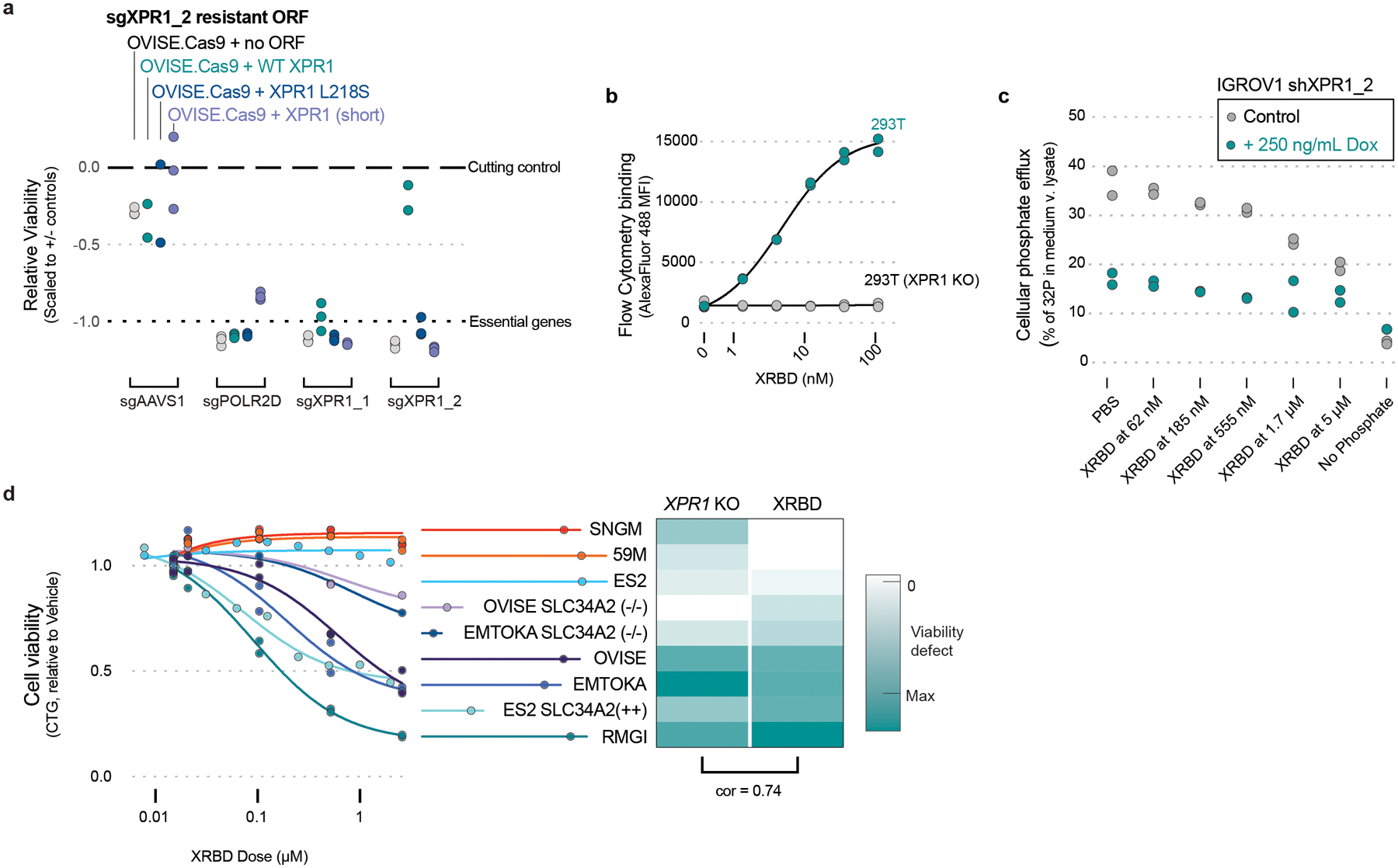
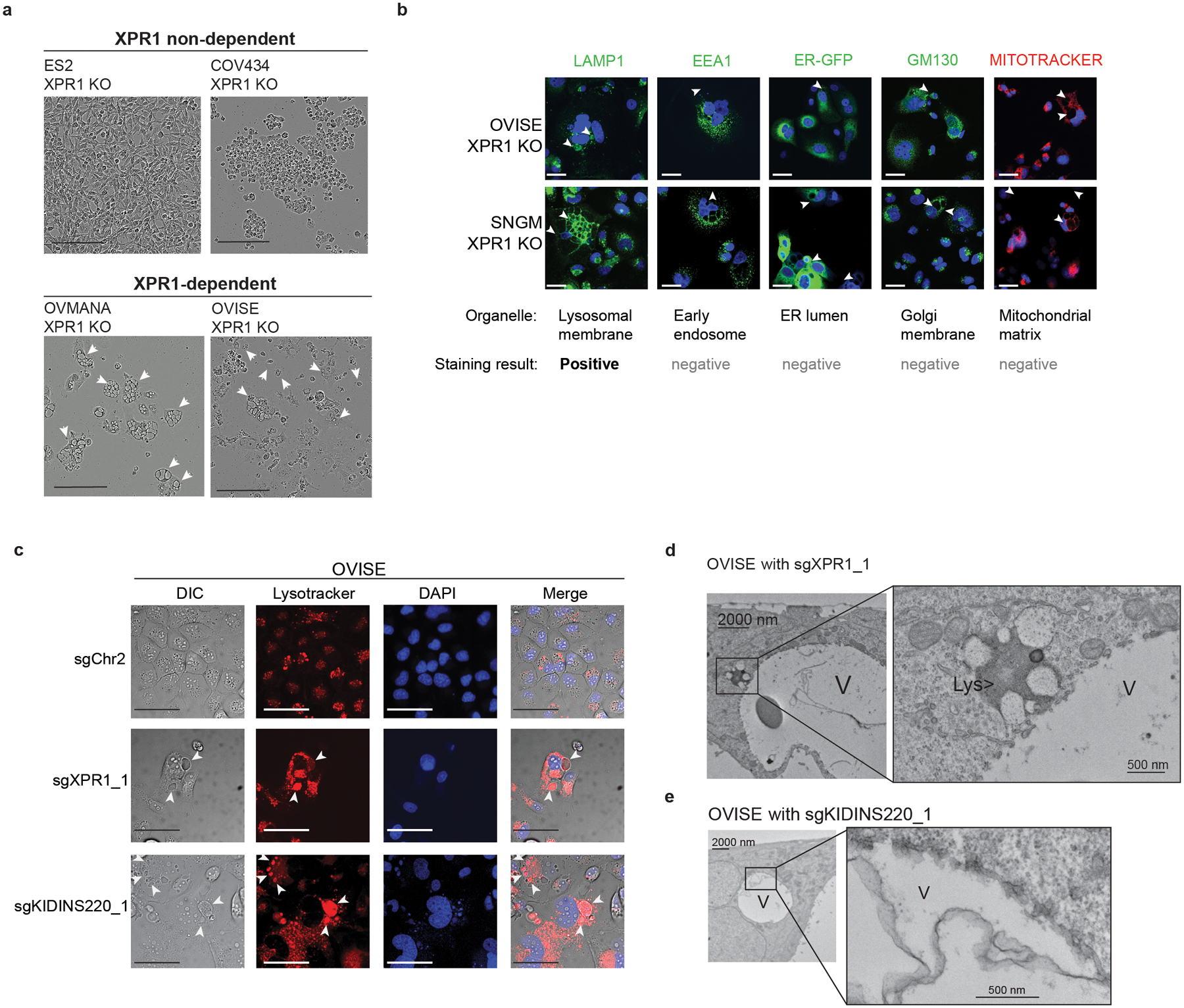
Despite advances in precision medicine, the clinical prospects for patients with ovarian and uterine cancers have not substantially improved. Here, we analyzed genome-scale CRISPR-Cas9 loss-of-function screens across 851 human cancer cell lines and found that frequent overexpression of SLC34A2-encoding a phosphate importer-is correlated with sensitivity to loss of the phosphate exporter XPR1, both in vitro and in vivo. In patient-derived tumor samples, we observed frequent PAX8-dependent overexpression of SLC34A2, XPR1 copy number amplifications and XPR1 messenger RNA overexpression. Mechanistically, in SLC34A2-high cancer cell lines, genetic or pharmacologic inhibition of XPR1-dependent phosphate efflux leads to the toxic accumulation of intracellular phosphate. Finally, we show that XPR1 requires the novel partner protein KIDINS220 for proper cellular localization and activity, and that disruption of this protein complex results in acidic "vacuolar" structures preceding cell death. These data point to the XPR1-KIDINS220 complex and phosphate dysregulation as a therapeutic vulnerability in ovarian cancer.
© 2022. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing Interests Statement
FV receives research funding from Novo Holdings. DER receives research funding from the Functional Genomics Consortium (Abbvie, Jannsen, Merck and Vir) and is a director of Addgene. TRG previously received cash and/or equity compensation for consulting to GlaxoSmithKline, Sherlock Biosciences and FORMA Therapeutics, currently is a paid consultant to Anji Pharmaceuticals, and receives research funding from Bayer HealthCare, Calico Life Sciences, and Novo Holdings. W.C.H. is a consultant for ThermoFisher, Solasta, MPM Capital, iTeos, Frontier Medicines, and Paraxel and is a Scientific Founder and serves on the Scientific Advisory Board (SAB) for KSQ Therapeutics. All other authors declare no competing interests.
Figures



References
-
- Lheureux S, Braunstein M & Oza AM Epithelial ovarian cancer: Evolution of management in the era of precision medicine. CA Cancer J. Clin 69, 280–304 (2019). - PubMed
-
- Sung H et al. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin 71, 209–249 (2021). - PubMed
-
- Boehm JS et al. Cancer research needs a better map. Nature 589, 514–516 (2021). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
