

High-fat diet disrupts REG3γ and gut microbial rhythms promoting metabolic dysfunction
- PMID: 35439436
- PMCID: PMC9281554
- DOI: 10.1016/j.chom.2022.03.030
High-fat diet disrupts REG3γ and gut microbial rhythms promoting metabolic dysfunction
Abstract
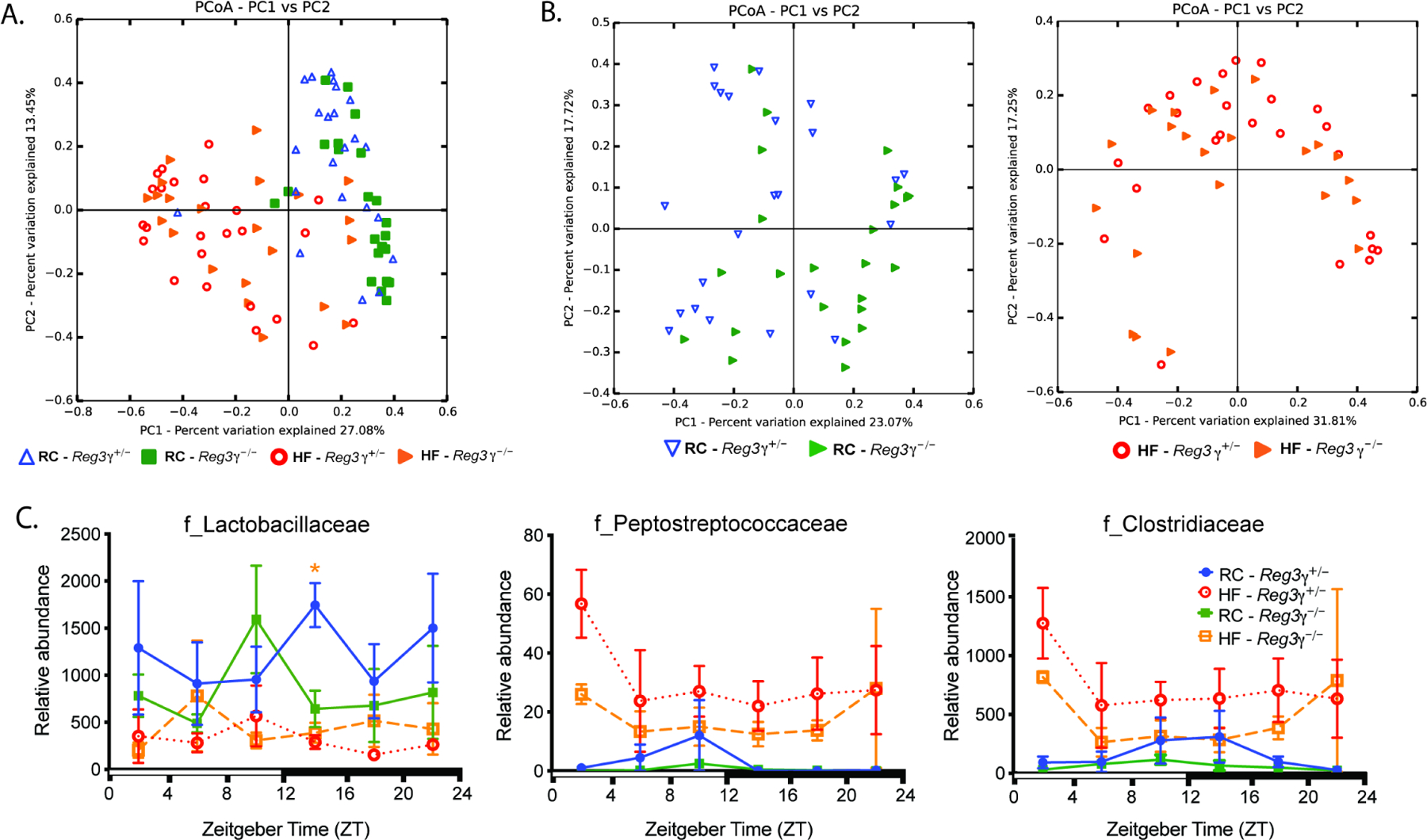
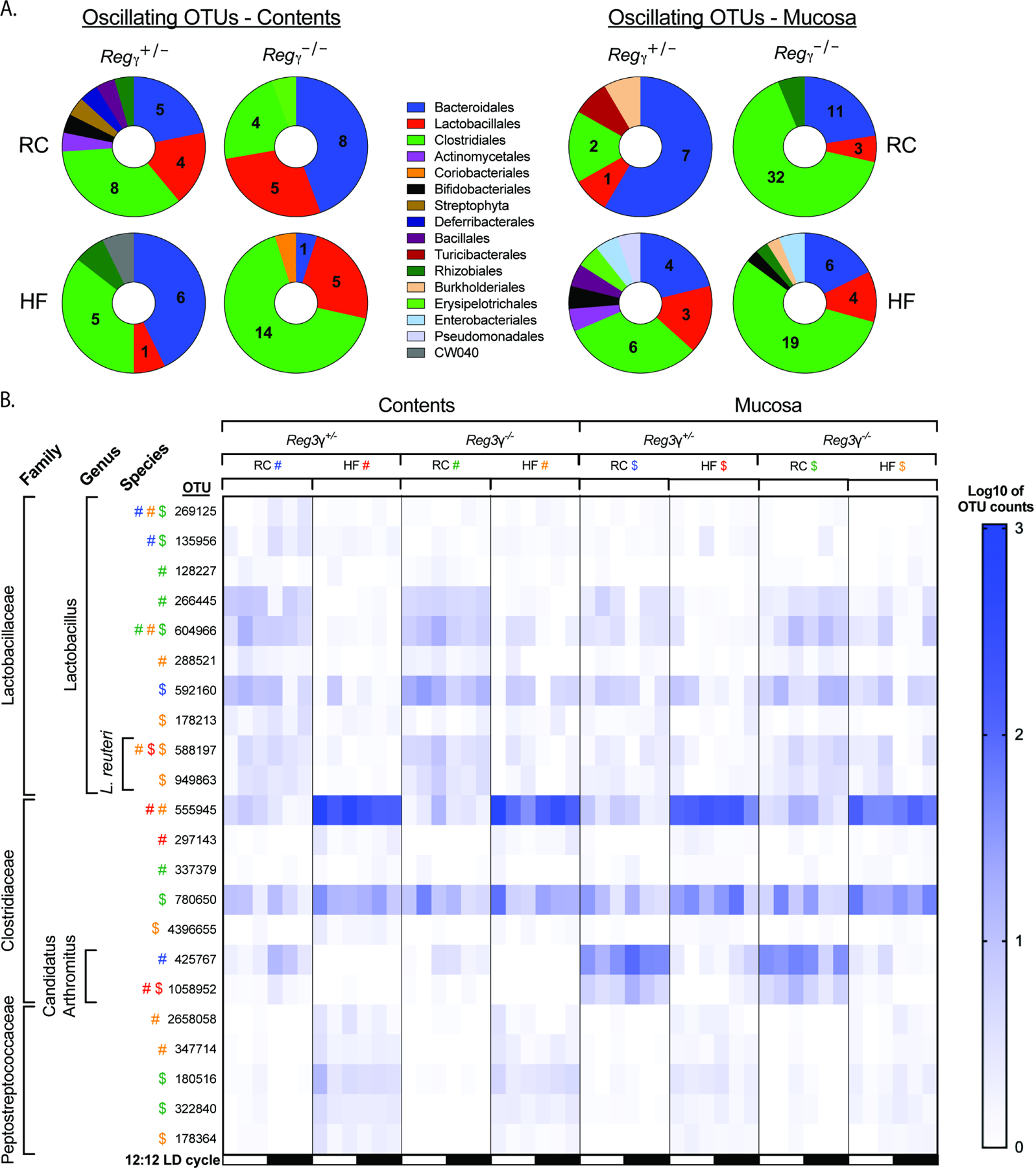
Gut microbial diurnal oscillations are important diet-dependent drivers of host circadian rhythms and metabolism ensuring optimal energy balance. However, the interplay between diet, microbes, and host factors sustaining intestinal oscillations is complex and poorly understood. Here, using a mouse model, we report the host C-type lectin antimicrobial peptide Reg3γ works with key ileal microbes to orchestrate these interactions in a bidirectional manner and does not correlate with the intestinal core circadian clock. High-fat diet is the primary driver of microbial oscillators that impair host metabolic homeostasis, resulting in arrhythmic host Reg3γ expression that secondarily drives abundance and oscillation of key gut microbes. This illustrates transkingdom coordination of biological rhythms primarily influenced by diet and reciprocal sensor-effector signals between host and microbial components, ultimately driving metabolism. Restoring the gut microbiota's capacity to sense dietary signals mediated by specific host factors such as Reg3γ could be harnessed to improve metabolic dysfunction.
Keywords: Reg3γ; circadian rhythms; diurnal oscillation; germ free; gut microbiota; high-fat diet; host-microbe interactions; innate immunity; organoid; small intestine.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.J.C. is a consultant for Thermo Fisher Scientific.
Figures
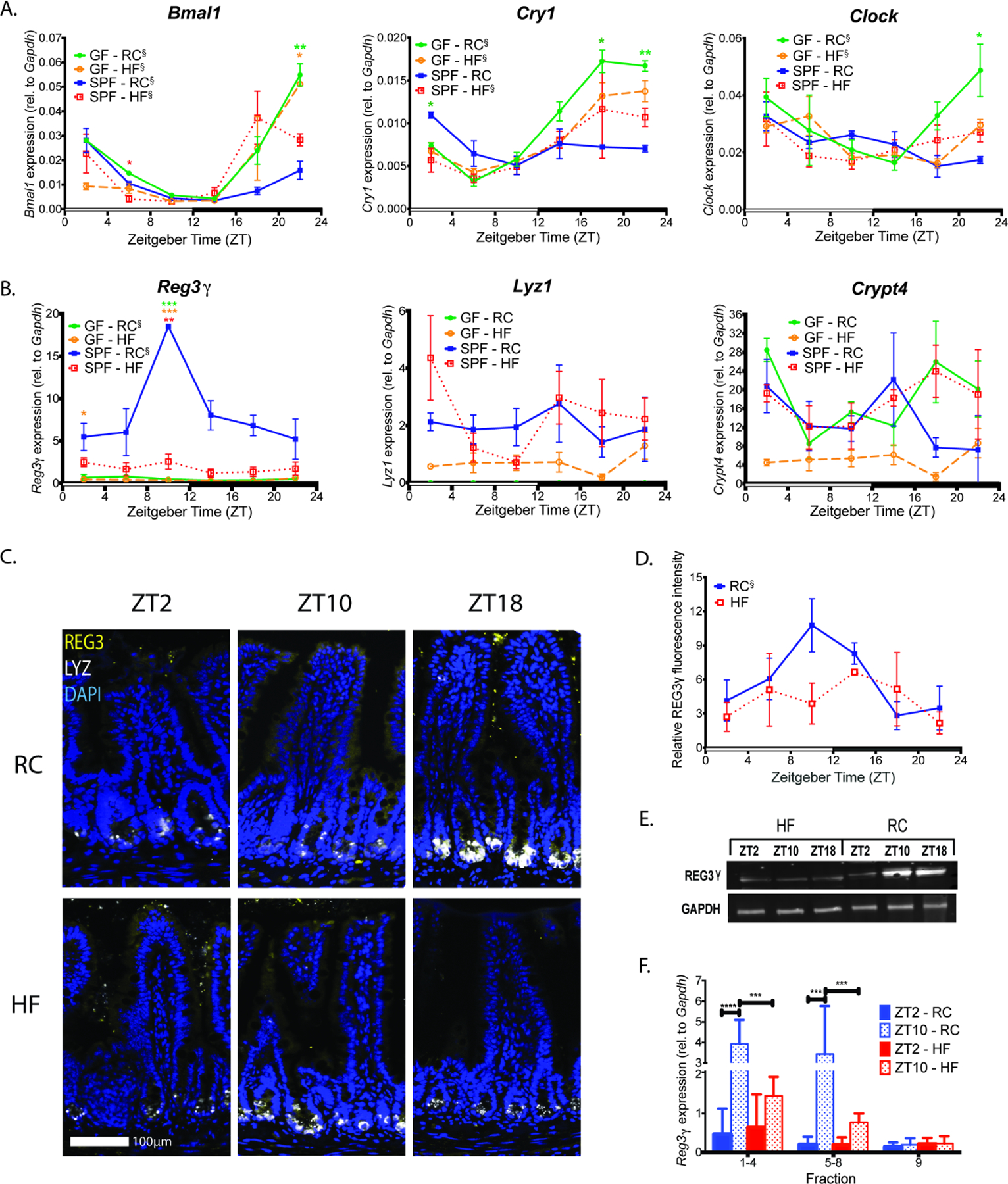
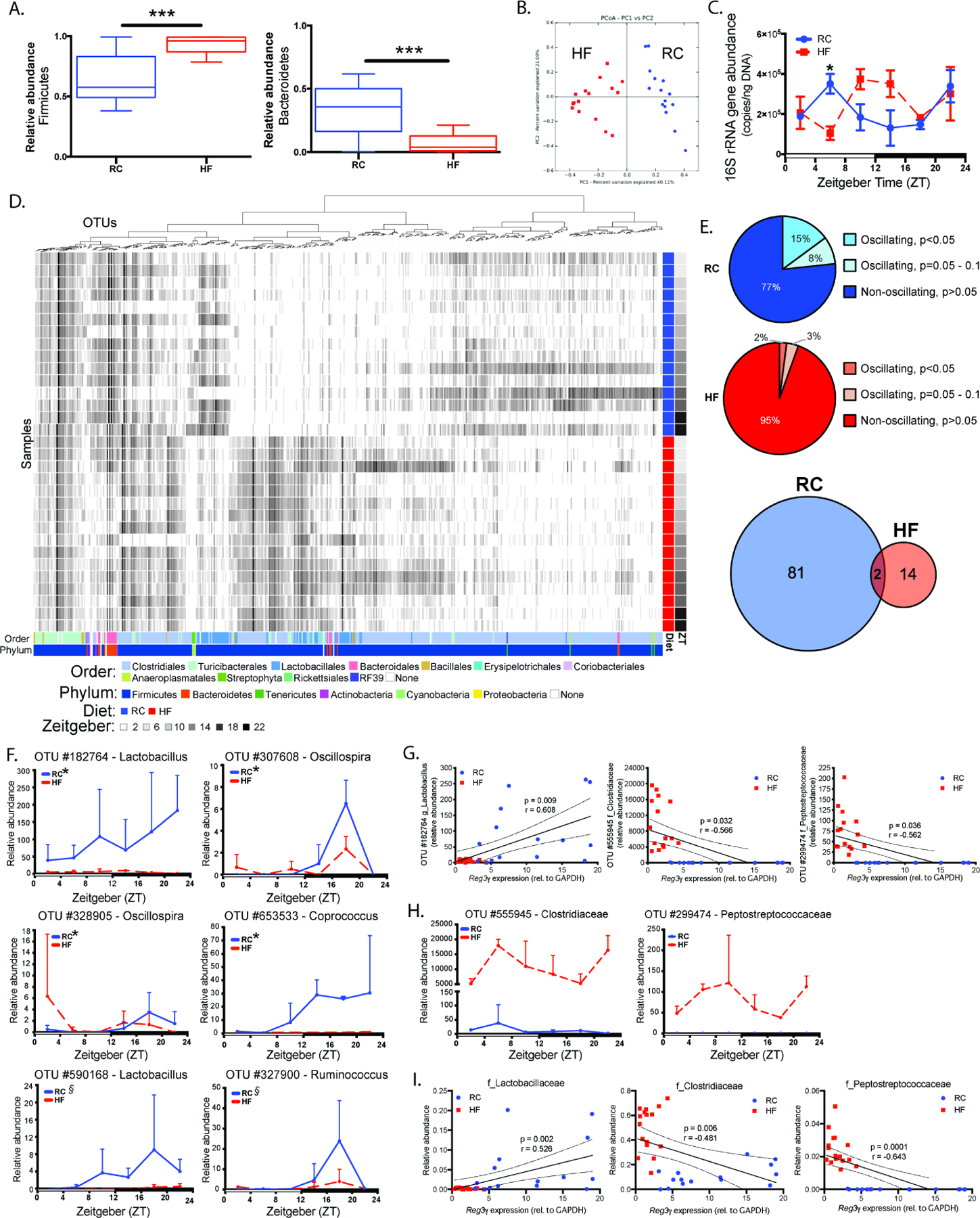
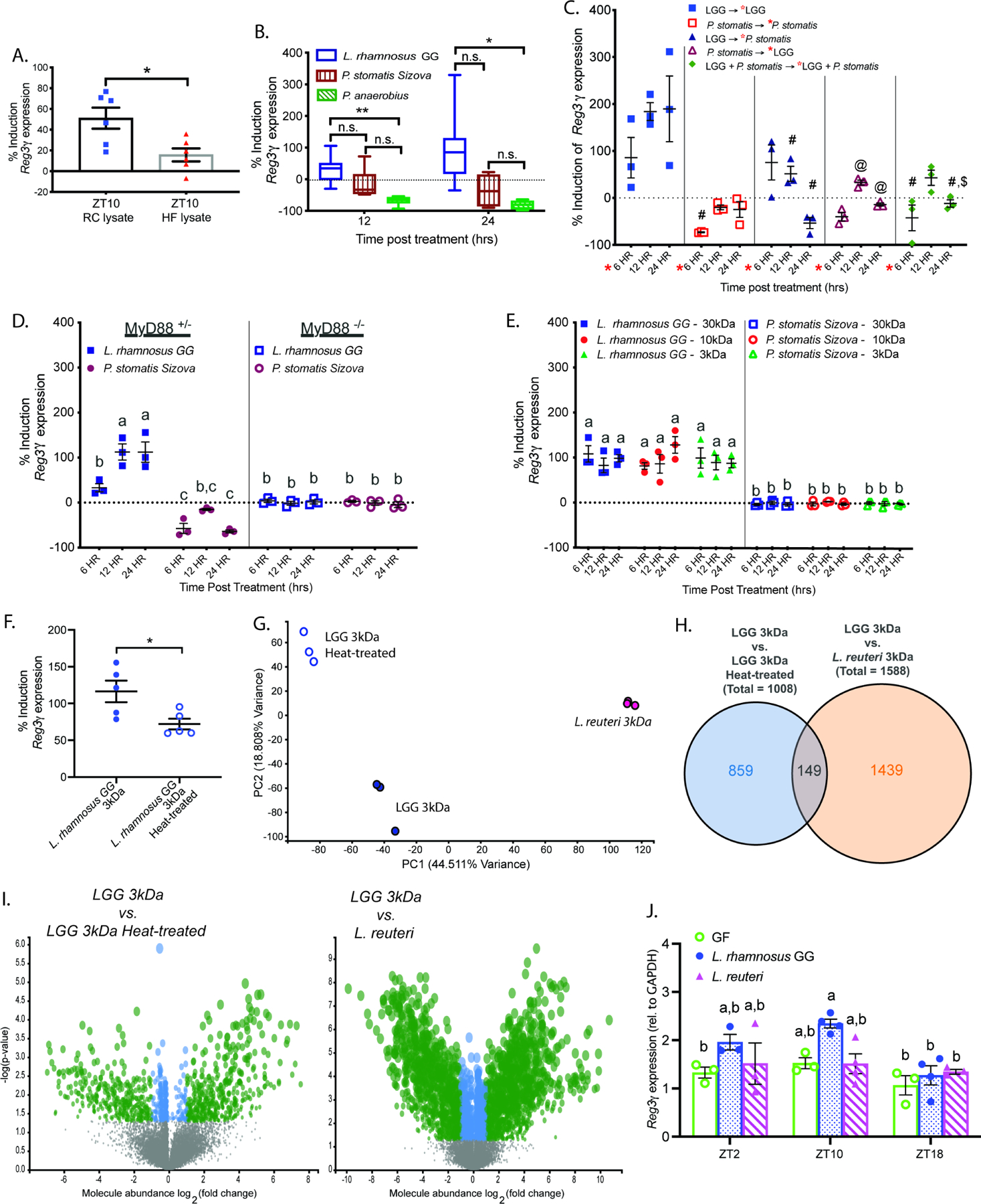
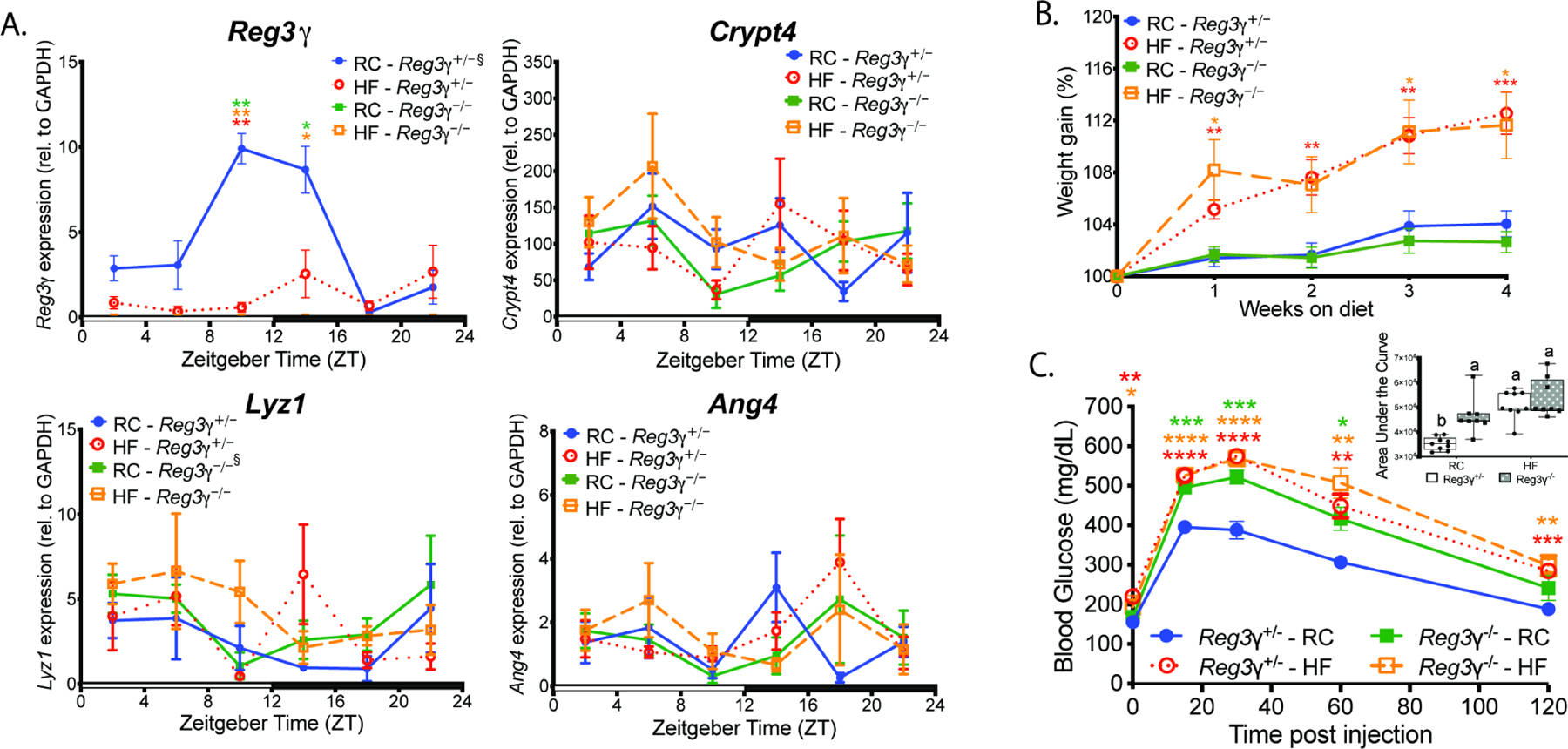
Comment on
-
Host-microbe circadian dynamics: Finding a rhythm and hitting a groove in scientific inquiry.Cell Host Microbe. 2022 Apr 13;30(4):458-462. doi: 10.1016/j.chom.2022.03.008. Cell Host Microbe. 2022. PMID: 35421343 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
