

Experimental demonstration and pan-structurome prediction of climate-associated riboSNitches in Arabidopsis
- PMID: 35440059
- PMCID: PMC9017077
- DOI: 10.1186/s13059-022-02656-4
Experimental demonstration and pan-structurome prediction of climate-associated riboSNitches in Arabidopsis
Abstract
Background: Genome-wide association studies (GWAS) aim to correlate phenotypic changes with genotypic variation. Upon transcription, single nucleotide variants (SNVs) may alter mRNA structure, with potential impacts on transcript stability, macromolecular interactions, and translation. However, plant genomes have not been assessed for the presence of these structure-altering polymorphisms or "riboSNitches."
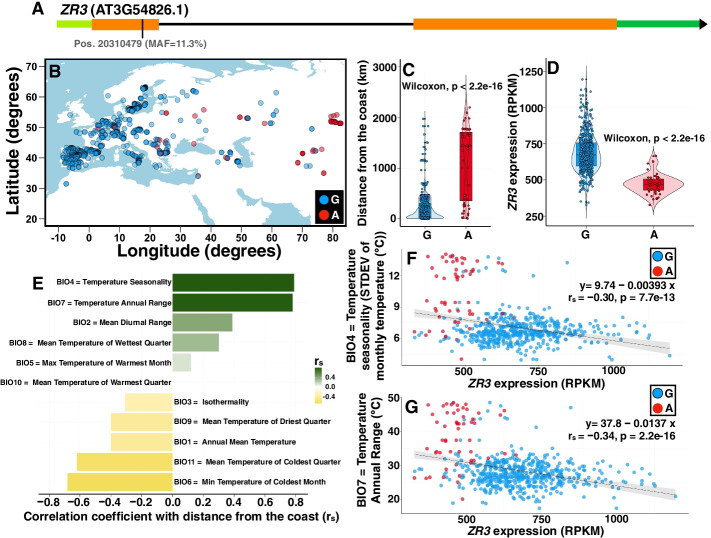
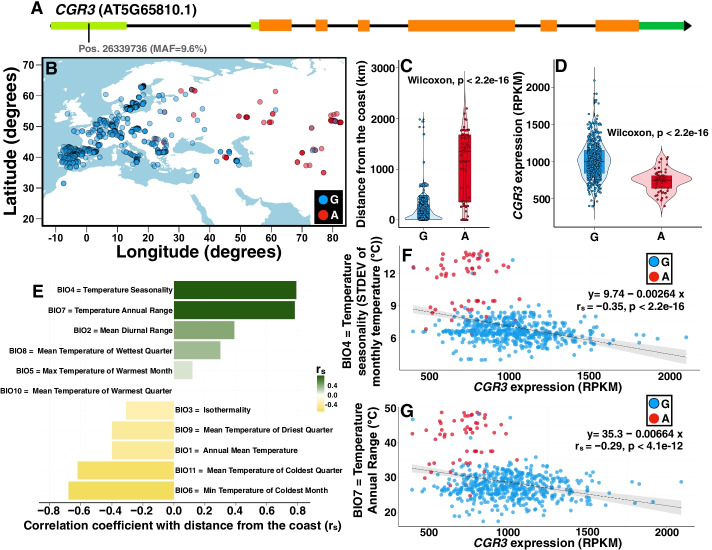
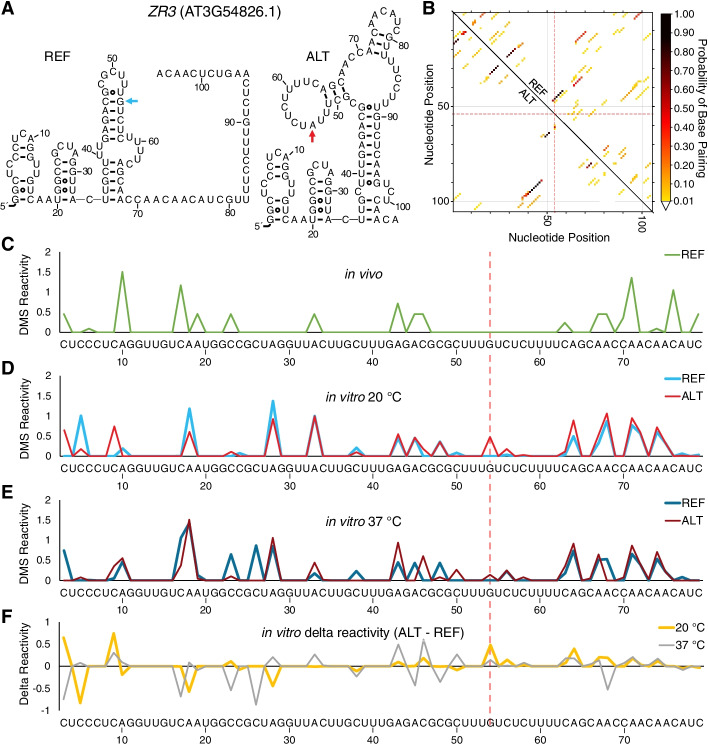
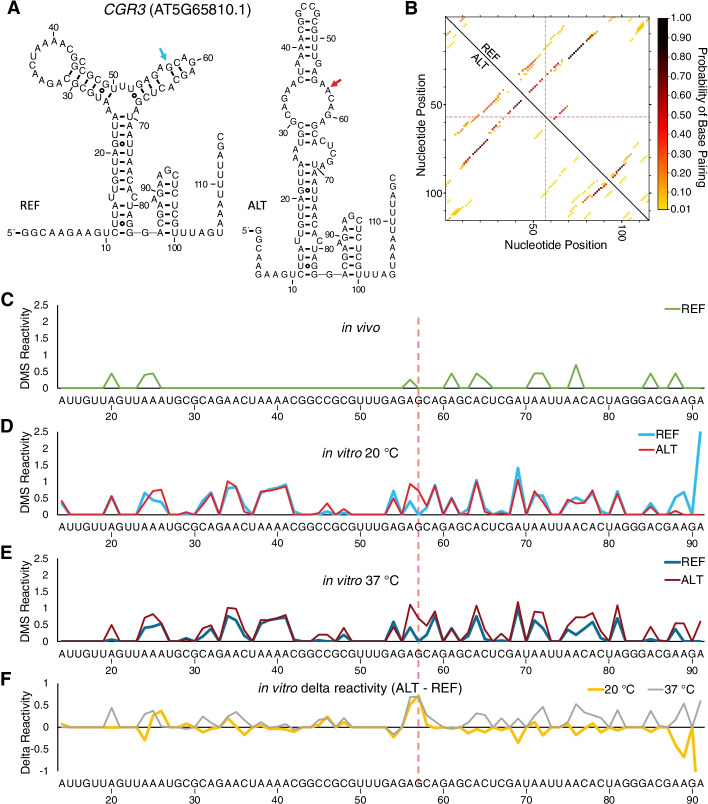
Results: We experimentally demonstrate the presence of riboSNitches in transcripts of two Arabidopsis genes, ZINC RIBBON 3 (ZR3) and COTTON GOLGI-RELATED 3 (CGR3), which are associated with continentality and temperature variation in the natural environment. These riboSNitches are also associated with differences in the abundance of their respective transcripts, implying a role in regulating the gene's expression in adaptation to local climate conditions. We then computationally predict riboSNitches transcriptome-wide in mRNAs of 879 naturally inbred Arabidopsis accessions. We characterize correlations between SNPs/riboSNitches in these accessions and 434 climate descriptors of their local environments, suggesting a role of these variants in local adaptation. We integrate this information in CLIMtools V2.0 and provide a new web resource, T-CLIM, that reveals associations between transcript abundance variation and local environmental variation.
Conclusion: We functionally validate two plant riboSNitches and, for the first time, demonstrate riboSNitch conditionality dependent on temperature, coining the term "conditional riboSNitch." We provide the first pan-genome-wide prediction of riboSNitches in plants. We expand our previous CLIMtools web resource with riboSNitch information and with 1868 additional Arabidopsis genomes and 269 additional climate conditions, which will greatly facilitate in silico studies of natural genetic variation, its phenotypic consequences, and its role in local adaptation.
Keywords: Arabidopsis thaliana; CLIMtools; Genome-wide association study (GWAS); Single nucleotide variant (SNV); Structurome; Transcriptome-wide association study (TWAS); riboSNitch.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
