

The role of procoagulant phospholipids on the surface of circulating blood cells in thrombosis and haemostasis
- PMID: 35440201
- PMCID: PMC9019515
- DOI: 10.1098/rsob.210318
The role of procoagulant phospholipids on the surface of circulating blood cells in thrombosis and haemostasis
Abstract
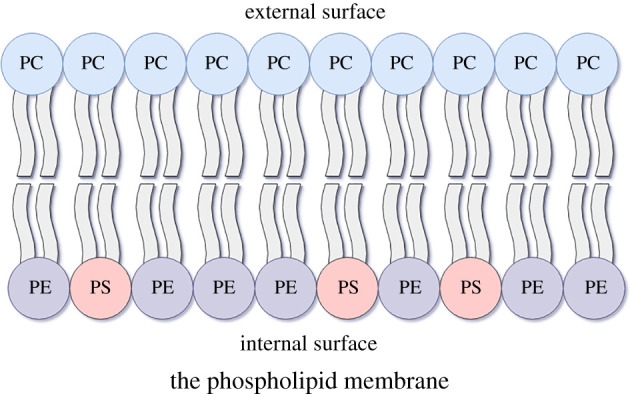
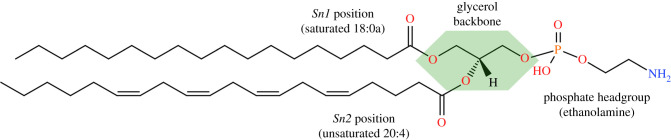
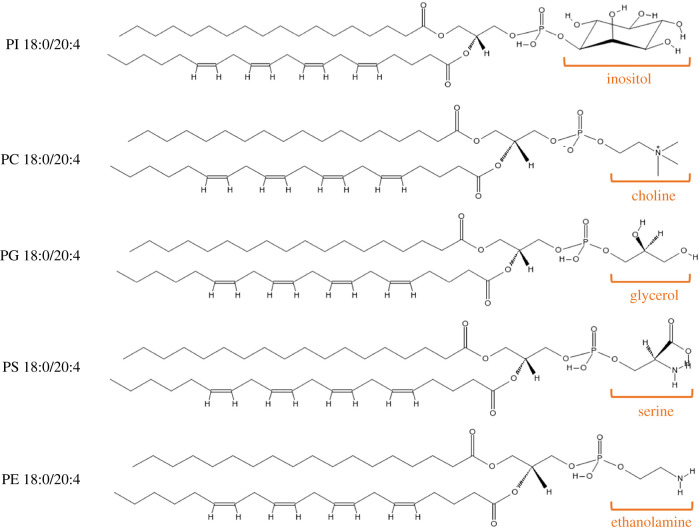
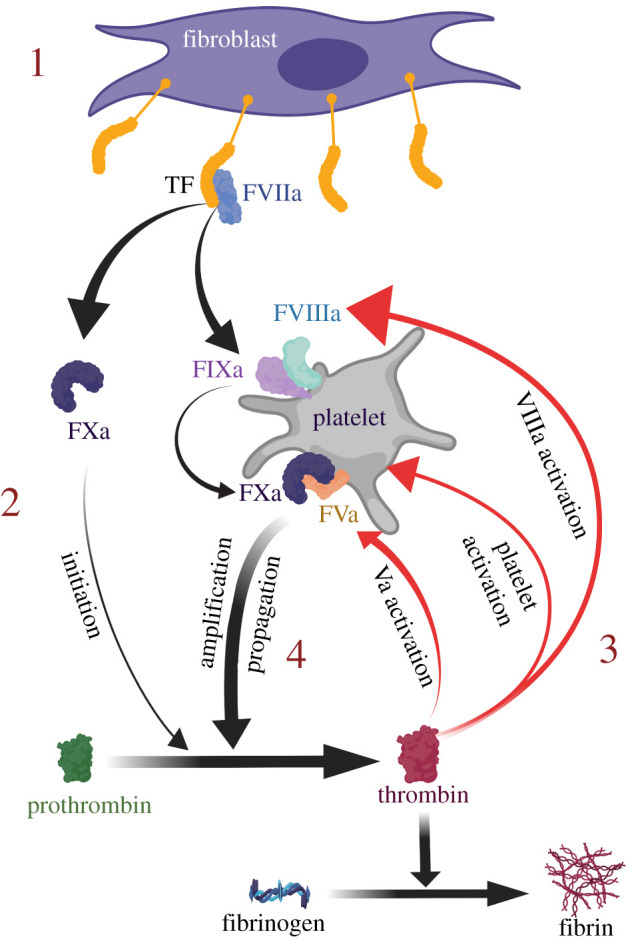
Phospholipids (PLs) are found in all cell types and are required for structural support and cell activation signalling pathways. In resting cells, PLs are asymmetrically distributed throughout the plasma membrane with native procoagulant aminophospholipids (aPLs) being actively maintained in the inner leaflet of the membrane. Upon platelet activation, aPLs rapidly externalize to the outer leaflet and are essential for supporting the coagulation cascade by providing binding sites for factors in the cell-based model. More recent work has uncovered a role for enzymatically oxidized PLs (eoxPLs) in facilitating coagulation, working in concert with native aPLs. Despite this, the role of aPLs and eoxPLs in thrombo-inflammatory conditions, such as arterial and venous thrombosis, has not been fully elucidated. In this review, we describe the biochemical structures, distribution and regulation of aPL externalization and summarize the literature on eoxPL generation in circulating blood cells. We focus on the currently understood role of these lipids in mediating coagulation reactions in vitro, in vivo and in human thrombotic disease. Finally, we highlight gaps in our understanding in how these lipids vary in health and disease, which may place them as future therapeutic targets for the management of thrombo-inflammatory conditions.
Keywords: coagulation; lipidomics; phospholipids; thrombosis.
Conflict of interest statement
All authors have no relevant affiliations or financial involvement with any organization or entity with a financial interest in or financial conflict with the subject matter or materials discussed in the manuscript.
Figures
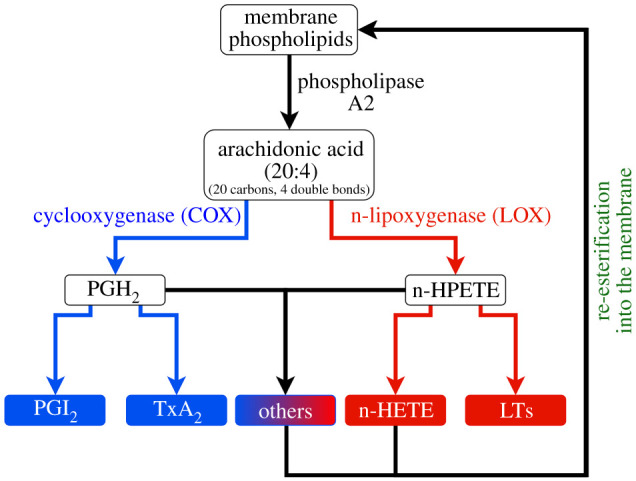
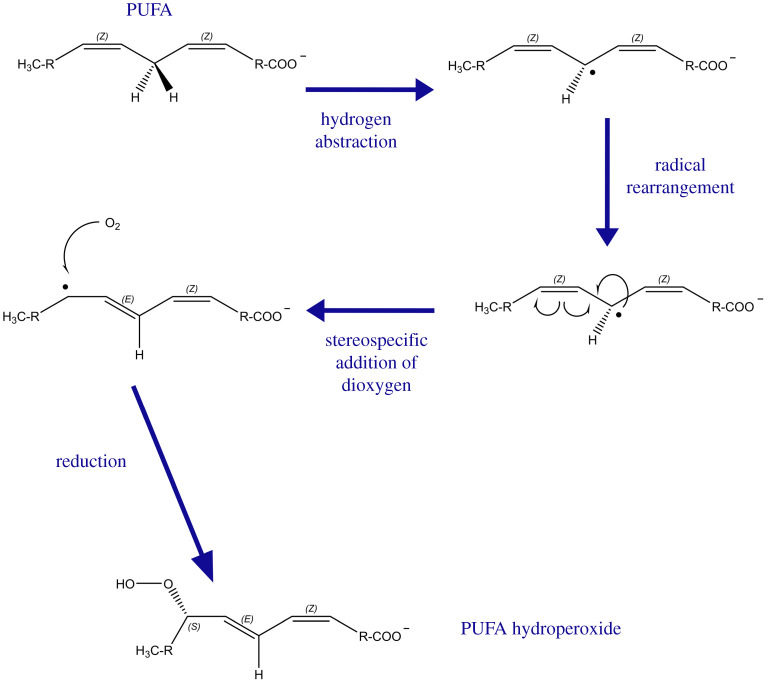
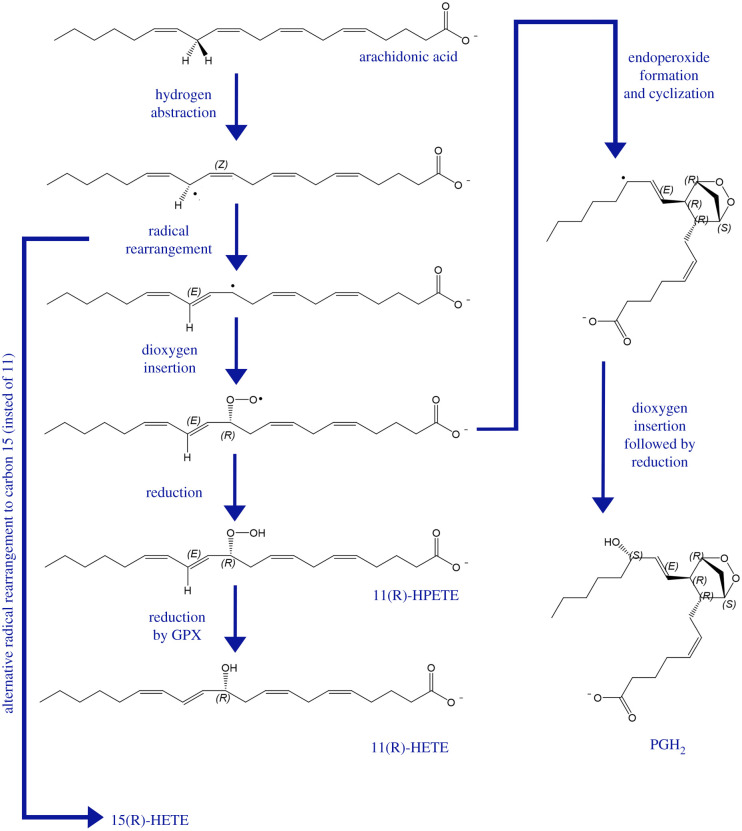
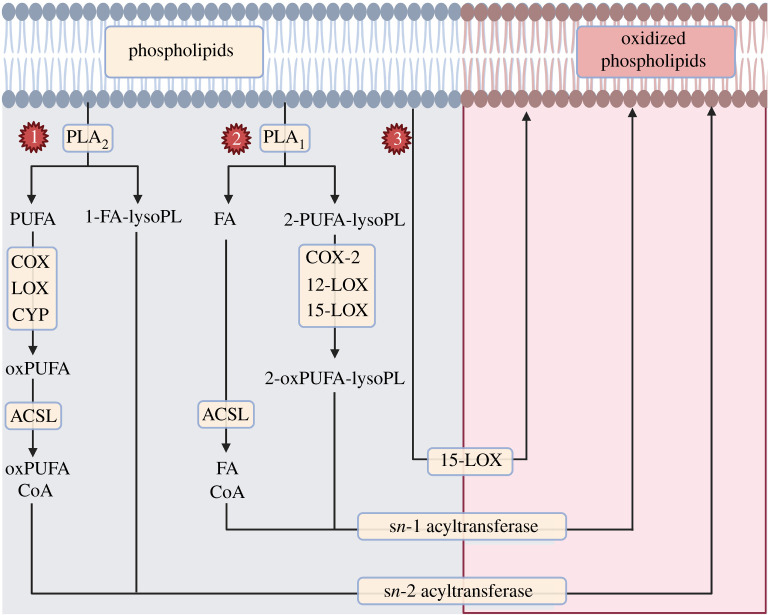
References
-
- Vance DE, Vance JE. 2008. Biochemistry of lipids, lipoproteins, and membranes, 5th edn. Amsterdam, The Netherlands: Elsevier.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical