

Secretory autophagy maintains proteostasis upon lysosome inhibition
- PMID: 35446347
- PMCID: PMC9036093
- DOI: 10.1083/jcb.202110151
Secretory autophagy maintains proteostasis upon lysosome inhibition
Abstract
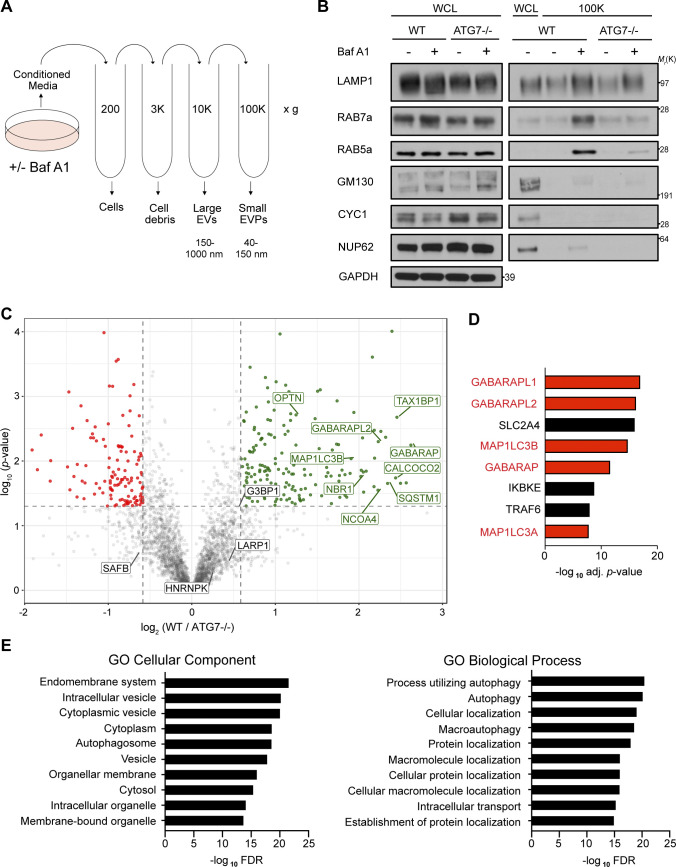
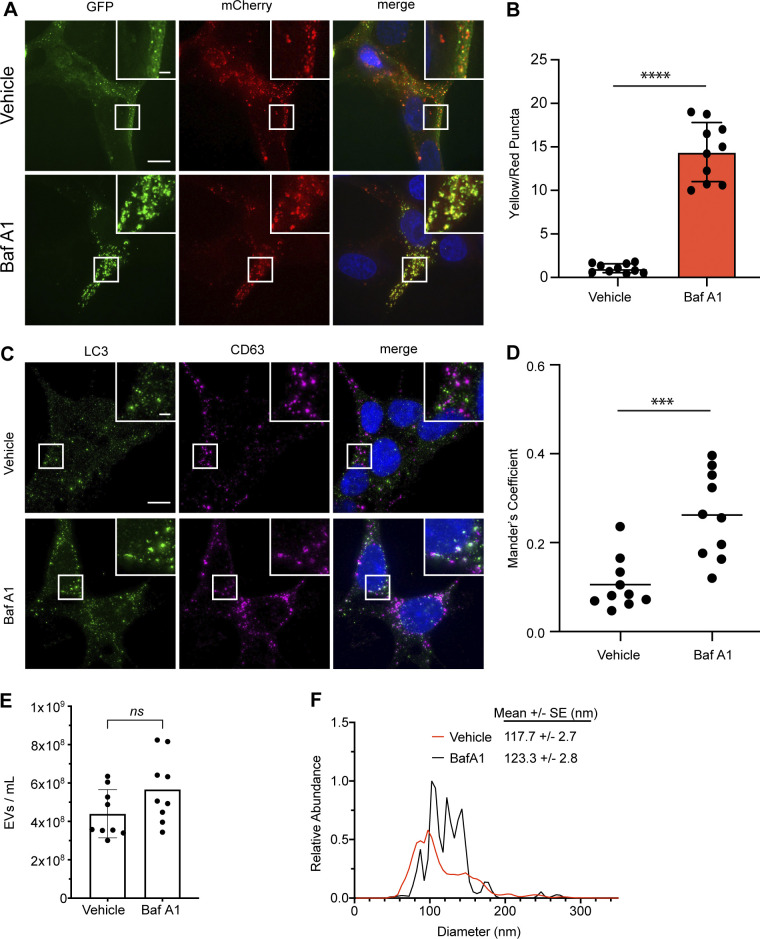
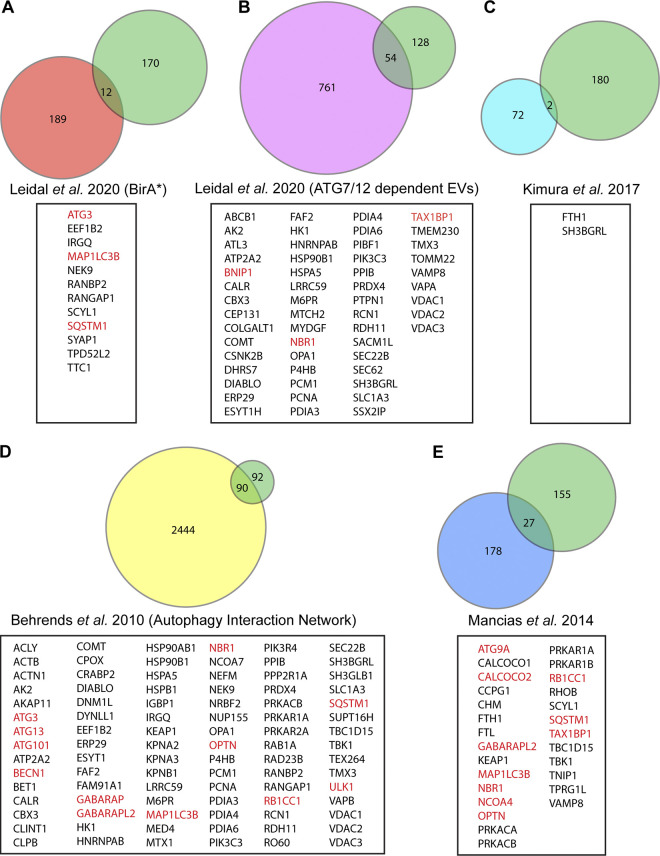
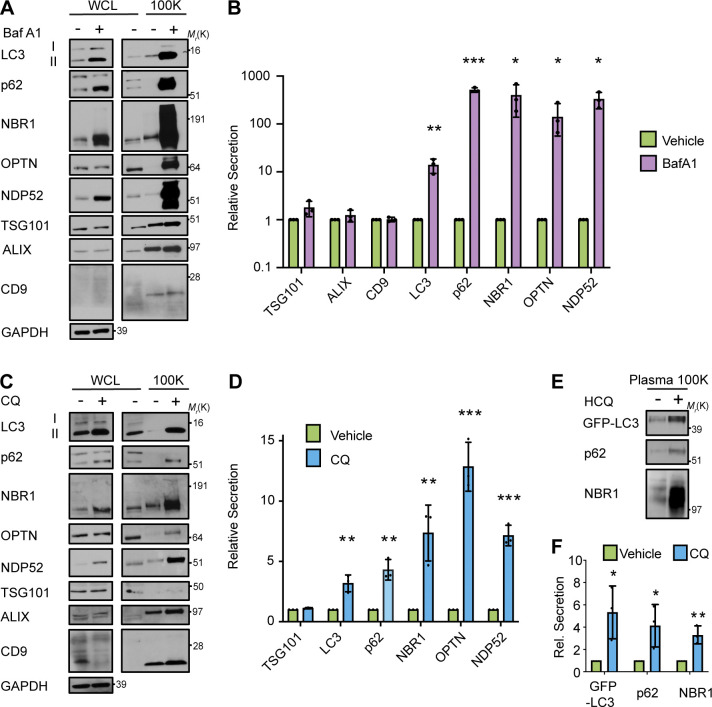
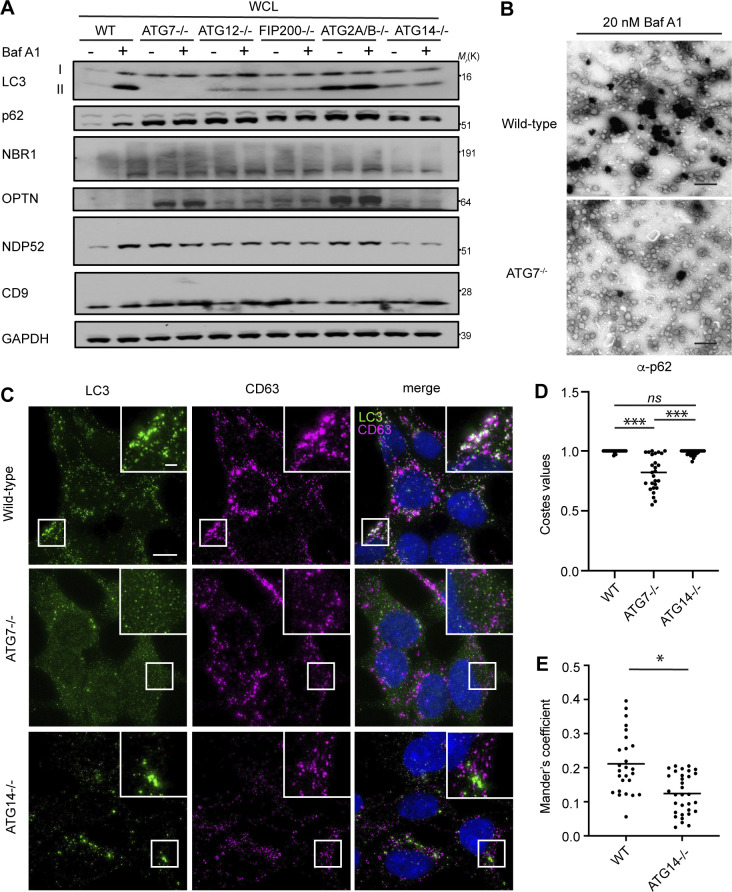
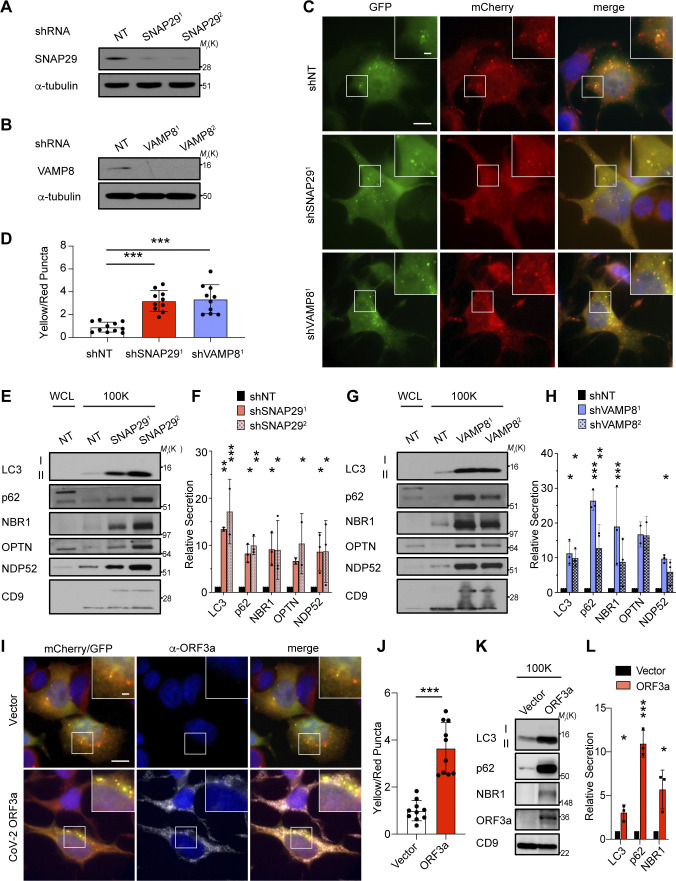
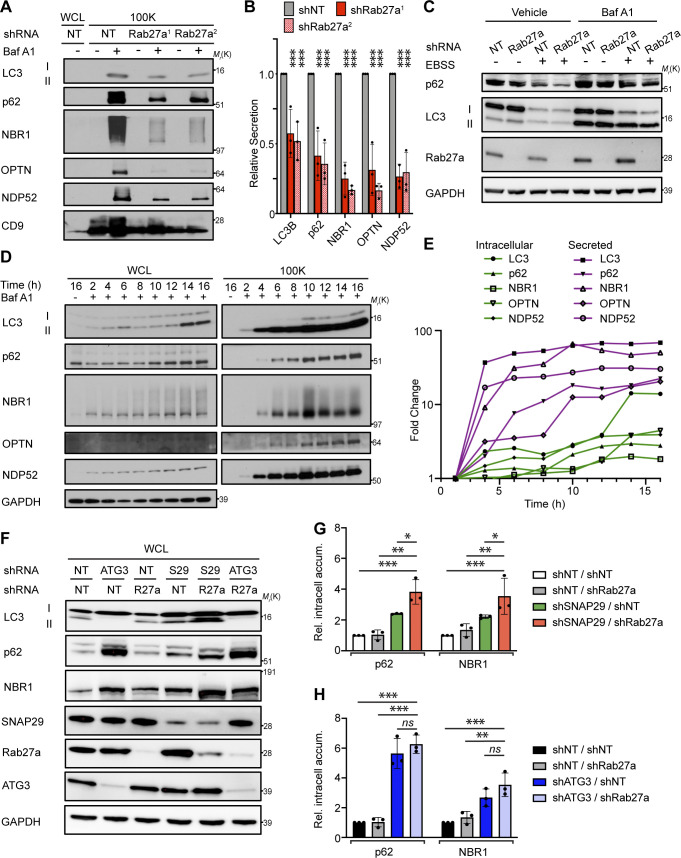
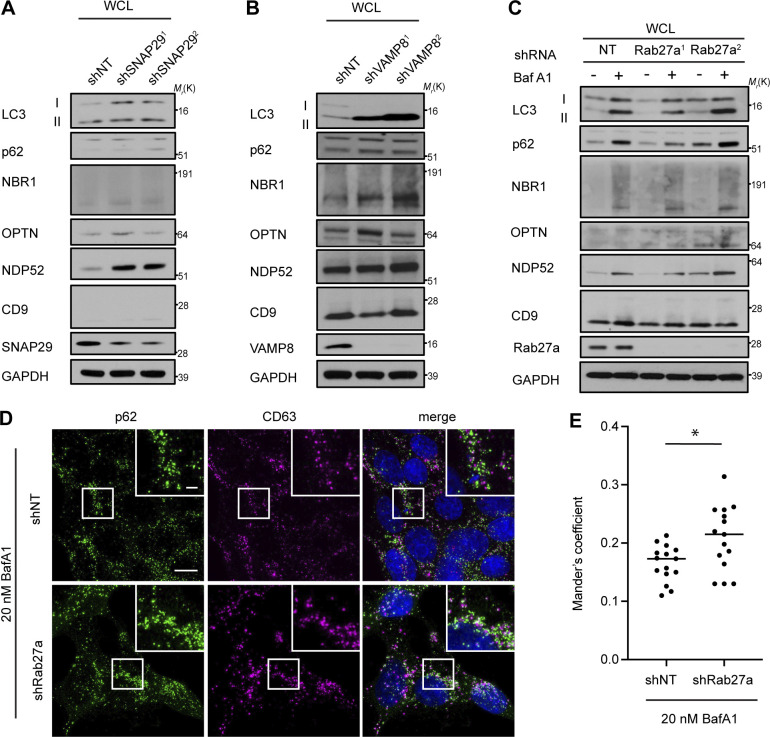
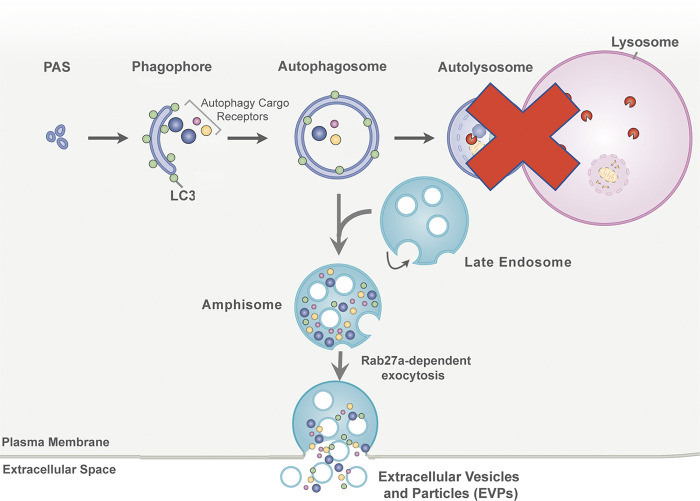
The endolysosome system plays central roles in both autophagic degradation and secretory pathways, including the release of extracellular vesicles and particles (EVPs). Although previous work reveals important interconnections between autophagy and EVP-mediated secretion, our understanding of these secretory events during endolysosome inhibition remains incomplete. Here, we delineate a secretory autophagy pathway upregulated in response to endolysosomal inhibition, which mediates EVP-associated release of autophagic cargo receptors, including p62/SQSTM1. This secretion is highly regulated and dependent on multiple ATGs required for autophagosome formation, as well as the small GTPase Rab27a. Furthermore, disrupting autophagosome maturation, either via genetic inhibition of autophagosome-to-autolysosome fusion or expression of SARS-CoV-2 ORF3a, is sufficient to induce EVP secretion of autophagy cargo receptors. Finally, ATG-dependent EVP secretion buffers against the intracellular accumulation of autophagy cargo receptors when classical autophagic degradation is impaired. Thus, we propose secretory autophagy via EVPs functions as an alternate route to clear sequestered material and maintain proteostasis during endolysosomal dysfunction or impaired autophagosome maturation.
© 2022 Solvik et al.
Figures
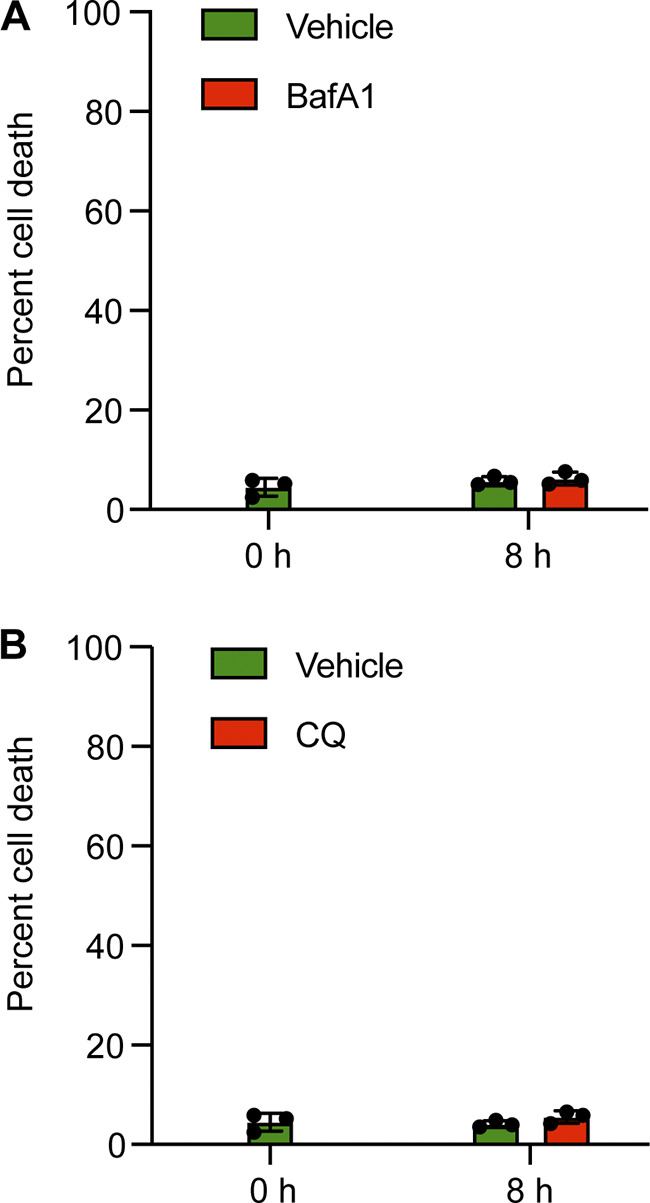
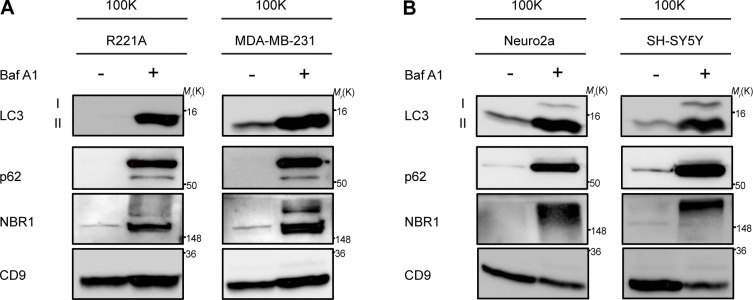
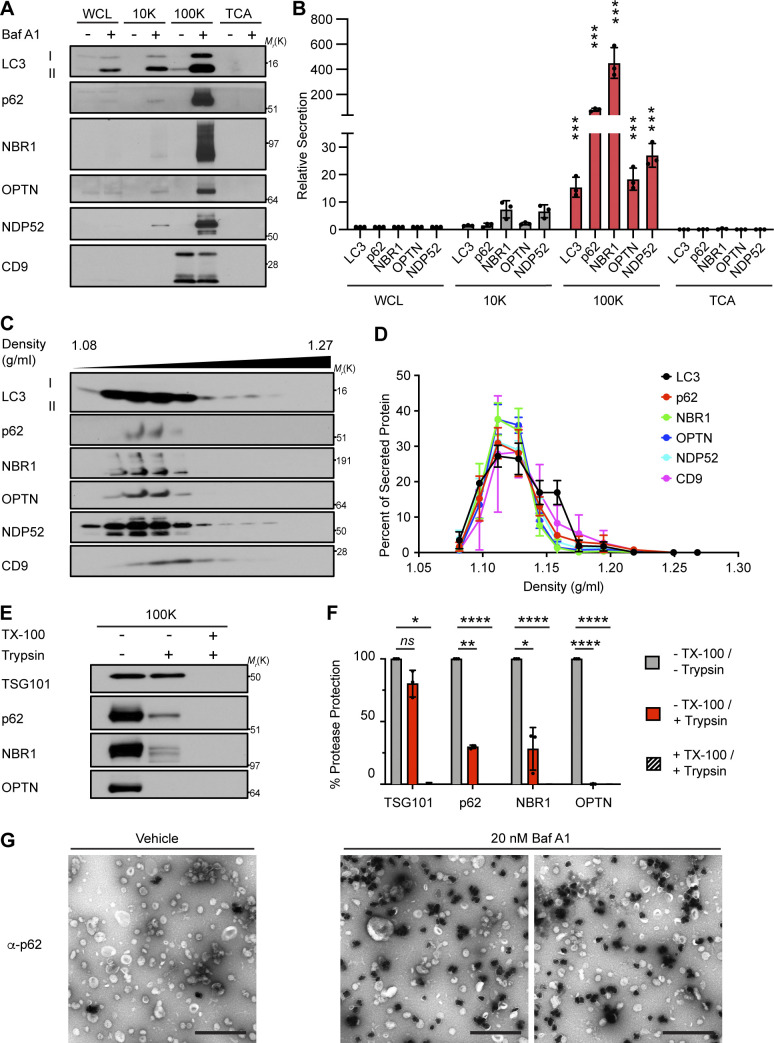
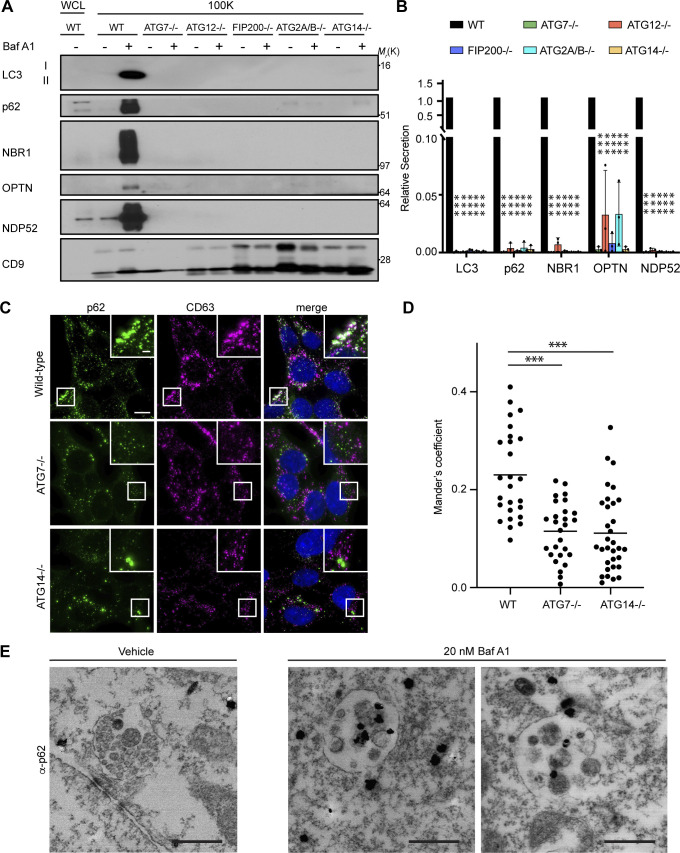
Comment in
-
Secretory autophagy during lysosome inhibition (SALI).Autophagy. 2022 Oct;18(10):2498-2499. doi: 10.1080/15548627.2022.2095788. Epub 2022 Jul 14. Autophagy. 2022. PMID: 35786367 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- Mark Foundation for Cancer Research
- Samuel Waxman Cancer Research Foundation
- R01 CA126792/CA/NCI NIH HHS/United States
- 201409BPF-335868/Banting Postdoctoral Fellowship
- R01 AG057462/AG/NIA NIH HHS/United States
- University of California, San Francisco
- 1650113/National Science Foundation
- F31 CA217015/CA/NCI NIH HHS/United States
- R01 CA226851/CA/NCI NIH HHS/United States
- 22805/Cancer Research Society
- R21 CA201849/CA/NCI NIH HHS/United States
- DFS 14-15/DRCRF/Damon Runyon Cancer Research Foundation/United States
- R01 CA213775/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
