

Cellular abundance of sodium phosphate cotransporter SLC20A1/PiT1 and phosphate uptake are controlled post-transcriptionally by ESCRT
- PMID: 35447110
- PMCID: PMC9123275
- DOI: 10.1016/j.jbc.2022.101945
Cellular abundance of sodium phosphate cotransporter SLC20A1/PiT1 and phosphate uptake are controlled post-transcriptionally by ESCRT
Abstract
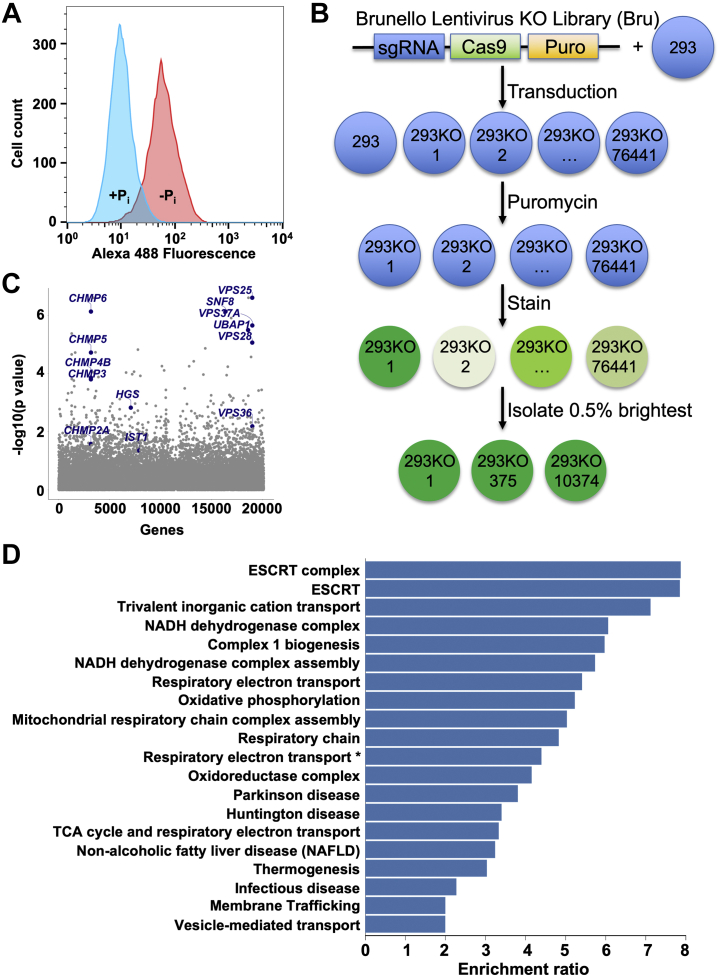
Inorganic phosphate is essential for human life. The widely expressed mammalian sodium/phosphate cotransporter SLC20A1/PiT1 mediates phosphate uptake into most cell types; however, while SLC20A1 is required for development, and elevated SLC20A1 expression is associated with vascular calcification and aggressive tumor growth, the mechanisms regulating SLC20A1 protein abundance are unknown. Here, we found that SLC20A1 protein expression is low in phosphate-replete cultured cells but is strikingly induced following phosphate starvation, whereas mRNA expression is high in phosphate-replete cells and only mildly increased by phosphate starvation. To identify regulators of SLC20A1 protein levels, we performed a genome-wide CRISPR-based loss-of-function genetic screen in phosphate-replete cells using SLC20A1 protein induction as readout. Our screen revealed that endosomal sorting complexes required for transport (ESCRT) machinery was essential for proper SLC20A1 protein downregulation in phosphate-replete cells. We show that SLC20A1 colocalizes with ESCRT and that ESCRT deficiency increases SLC20A1 protein and phosphate uptake into cells. We also found numerous additional candidate regulators of mammalian phosphate homeostasis, including genes modifying protein ubiquitination and the Krebs cycle and oxidative phosphorylation pathways. Many of these targets have not been previously implicated in this process. We present here a model in which SLC20A1 protein abundance and phosphate uptake are tonically negatively regulated post-transcriptionally in phosphate-replete cells through direct ESCRT-mediated SLC20A1 degradation. Moreover, our screening results provide a comprehensive resource for future studies to elucidate the mechanisms governing cellular phosphate homeostasis. We conclude that genome-wide CRISPR-based genetic screening is a powerful tool to discover proteins and pathways relevant to physiological processes.
Keywords: CRISPR/Cas9; cell metabolism; cell surface protein; endosomal sorting complexes required for transport; genome-wide forward genetic screen; membrane transport; phosphate transporter; protein degradation.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures





References
-
- Goldford J.E., Hartman H., Smith T.F., Segre D. Remnants of an ancient metabolism without phosphate. Cell. 2017;168:1126–1134.e9. - PubMed
-
- Krebs E.G., Graves J.D. Interactions between protein kinases and proteases in cellular signaling and regulation. Adv. Enzyme Regul. 2000;40:441–470. - PubMed
-
- Vance J.E. Phospholipid synthesis and transport in mammalian cells. Traffic. 2015;16:1–18. - PubMed
-
- Florenzano P., Cipriani C., Roszko K.L., Fukumoto S., Collins M.T., Minisola S., Pepe J. Approach to patients with hypophosphataemia. Lancet Diabetes Endocrinol. 2020;8:163–174. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
