

Calpain-mediated proteolytic production of free amino acids in vascular endothelial cells augments obesity-induced hepatic steatosis
- PMID: 35447117
- PMCID: PMC9110893
- DOI: 10.1016/j.jbc.2022.101953
Calpain-mediated proteolytic production of free amino acids in vascular endothelial cells augments obesity-induced hepatic steatosis
Abstract
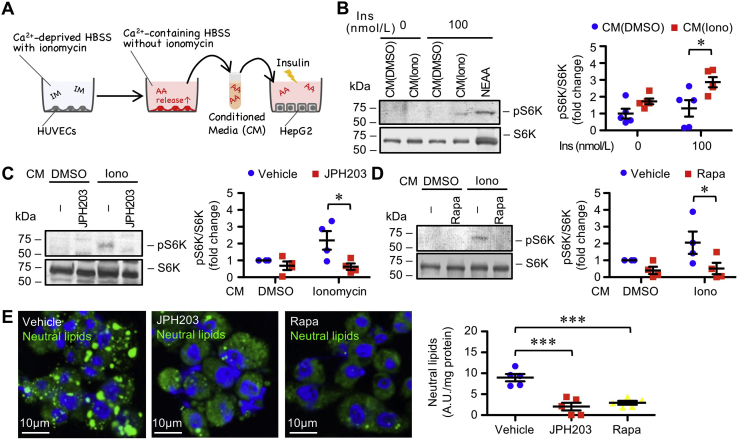
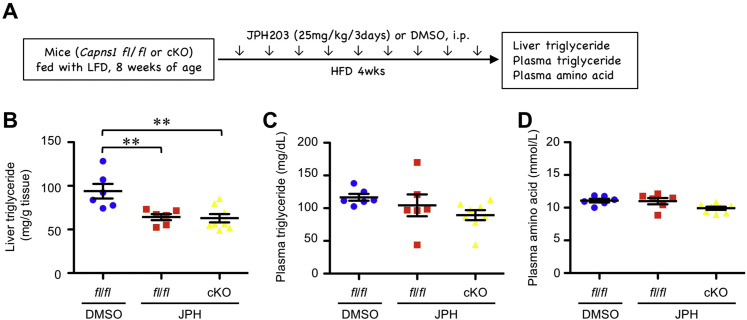
Free amino acids that accumulate in the plasma of patients with diabetes and obesity influence lipid metabolism and protein synthesis in the liver. The stress-inducible intracellular protease calpain proteolyzes various substrates in vascular endothelial cells (ECs), although its contribution to the supply of free amino acids in the liver microenvironment remains enigmatic. In the present study, we showed that calpains are associated with free amino acid production in cultured ECs. Furthermore, conditioned media derived from calpain-activated ECs facilitated the phosphorylation of ribosomal protein S6 kinase (S6K) and de novo lipogenesis in hepatocytes, which were abolished by the amino acid transporter inhibitor, JPH203, and the mammalian target of rapamycin complex 1 inhibitor, rapamycin. Meanwhile, calpain-overexpressing capillary-like ECs were observed in the livers of high-fat diet-fed mice. Conditional KO of EC/hematopoietic Capns1, which encodes a calpain regulatory subunit, diminished levels of branched-chain amino acids in the hepatic microenvironment without altering plasma amino acid levels. Concomitantly, conditional KO of Capns1 mitigated hepatic steatosis without normalizing body weight and the plasma lipoprotein profile in an amino acid transporter-dependent manner. Mice with targeted Capns1 KO exhibited reduced phosphorylation of S6K and maturation of lipogenic factor sterol regulatory element-binding protein 1 in hepatocytes. Finally, we show that bone marrow transplantation negated the contribution of hematopoietic calpain systems. We conclude that overactivation of calpain systems may be responsible for the production of free amino acids in ECs, which may be sufficient to potentiate S6K/sterol regulatory element-binding protein 1-induced lipogenesis in surrounding hepatocytes.
Keywords: calpastatin; capillarization; diabetes; inflammation; nonalcoholic fatty liver disease; pathological angiogenesis; sinusoidal liver endothelial cells; steatohepatitis; triglyceride; ubiquitin.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures





References
-
- Chalasani N., Younossi Z., Lavine J.E., Diehl A.M., Brunt E.M., Cusi K., Charlton M., Sanyal A.J. The diagnosis and management of non-alcoholic fatty liver disease: Practice guideline by the American gastroenterological association, American association for the study of liver diseases, and American college of gastroenterology. Gastroenterology. 2012;142:1592–1609. - PubMed
-
- Anderson N., Borlak J. Molecular mechanisms and therapeutic targets in steatosis and steatohepatitis. Pharmacol. Rev. 2008;60:311–357. - PubMed
-
- Le Couteur D.G., Solon-Biet S.M., Cogger V.C., Ribeiro R., de Cabo R., Raubenheimer D., Cooney G.J., Simpson S.J. Branched chain amino acids, aging and age-related health. Ageing Res. Rev. 2020;64:101198. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
