

Enantiomeric Discrimination in Insects: The Role of OBPs and ORs
- PMID: 35447810
- PMCID: PMC9030700
- DOI: 10.3390/insects13040368
Enantiomeric Discrimination in Insects: The Role of OBPs and ORs
Abstract
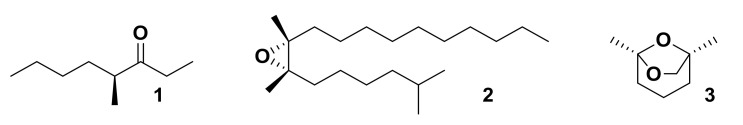
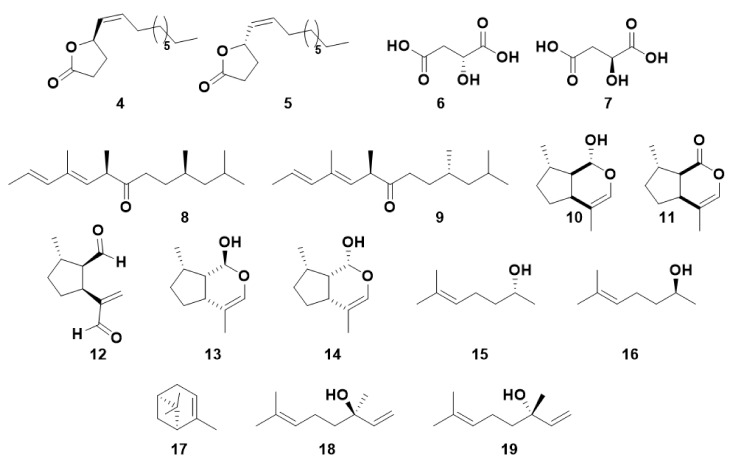
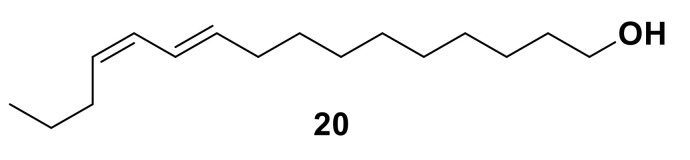
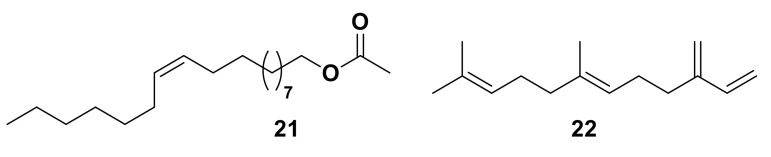
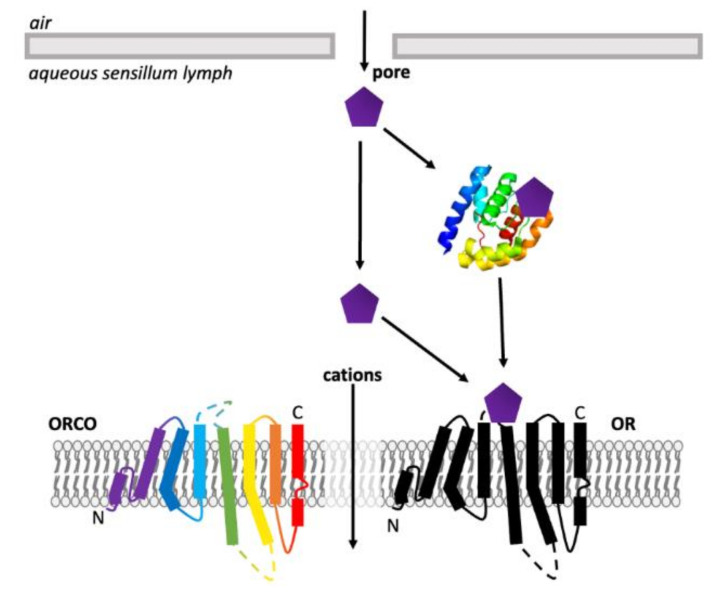
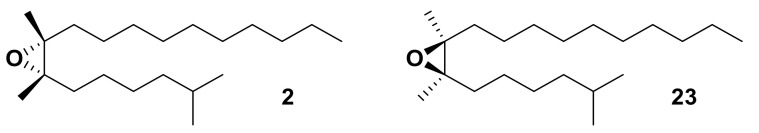
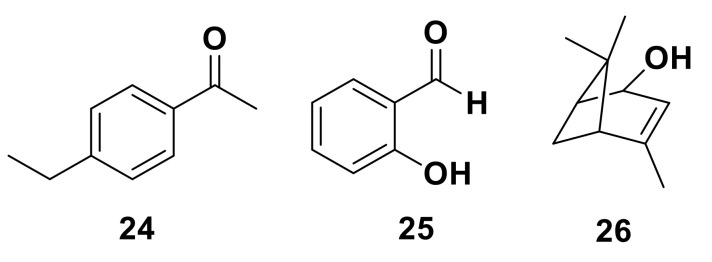
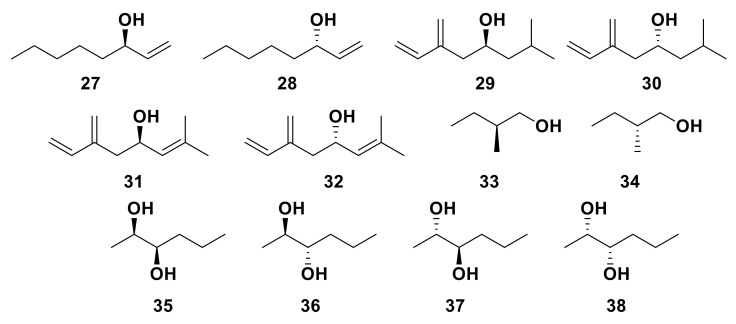
Olfaction is a complex recognition process that is critical for chemical communication in insects. Though some insect species are capable of discrimination between compounds that are structurally similar, little is understood about how this high level of discrimination arises. Some insects rely on discriminating between enantiomers of a compound, demonstrating an ability for highly selective recognition. The role of two major peripheral olfactory proteins in insect olfaction, i.e., odorant-binding proteins (OBPs) and odorant receptors (ORs) has been extensively studied. OBPs and ORs have variable discrimination capabilities, with some found to display highly specialized binding capability, whilst others exhibit promiscuous binding activity. A deeper understanding of how odorant-protein interactions induce a response in an insect relies on further analysis such as structural studies. In this review, we explore the potential role of OBPs and ORs in highly specific recognition, specifically enantiomeric discrimination. We summarize the state of research into OBP and OR function and focus on reported examples in the literature of clear enantiomeric discrimination by these proteins.
Keywords: chemical ecology; chemosensory; chiral; enantiomeric discrimination; insect; odorant receptors; odorant-binding proteins; olfaction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
