

The ATG5 interactome links clathrin-mediated vesicular trafficking with the autophagosome assembly machinery
- PMID: 35449600
- PMCID: PMC9015699
- DOI: 10.1080/27694127.2022.2042054
The ATG5 interactome links clathrin-mediated vesicular trafficking with the autophagosome assembly machinery
Abstract
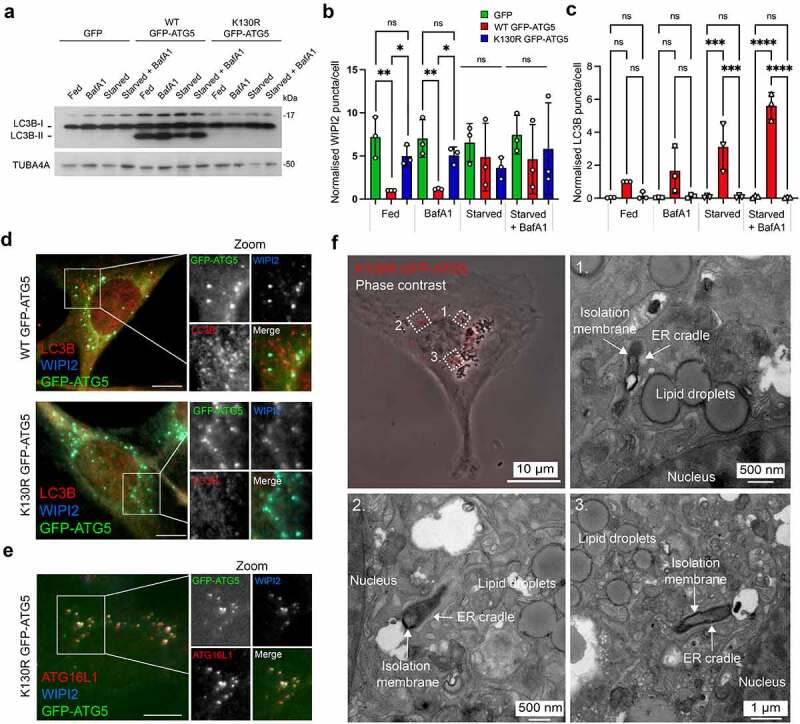
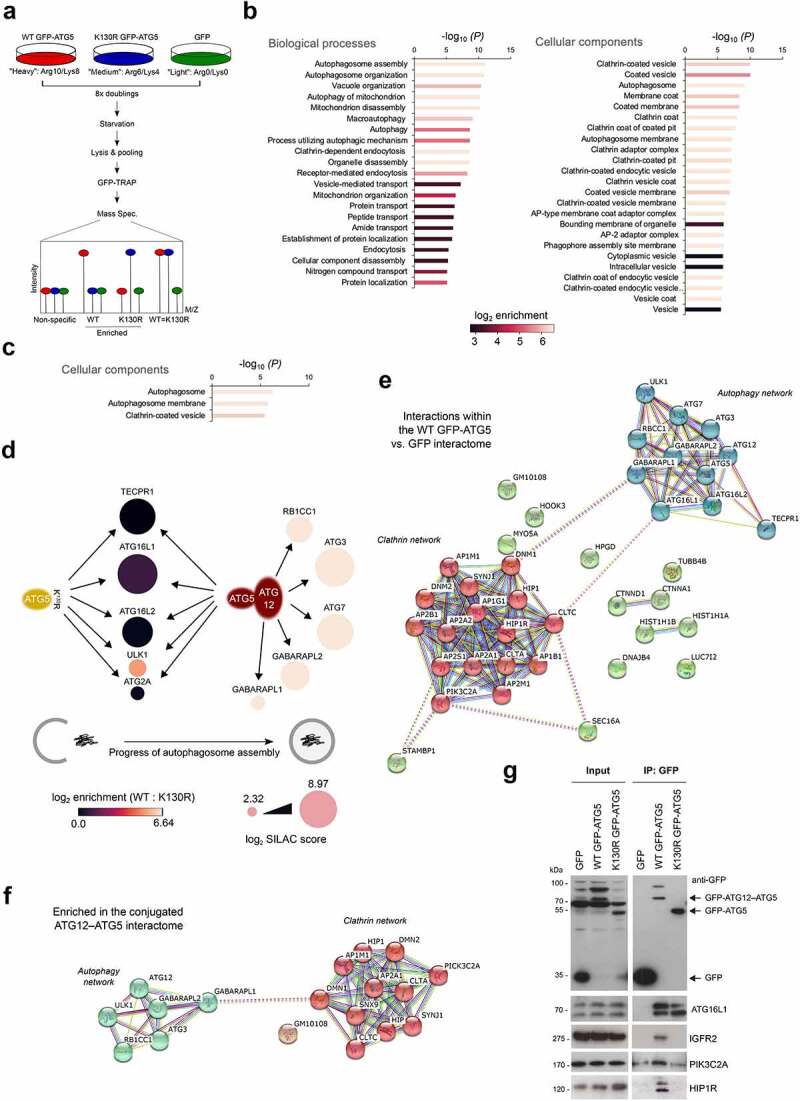
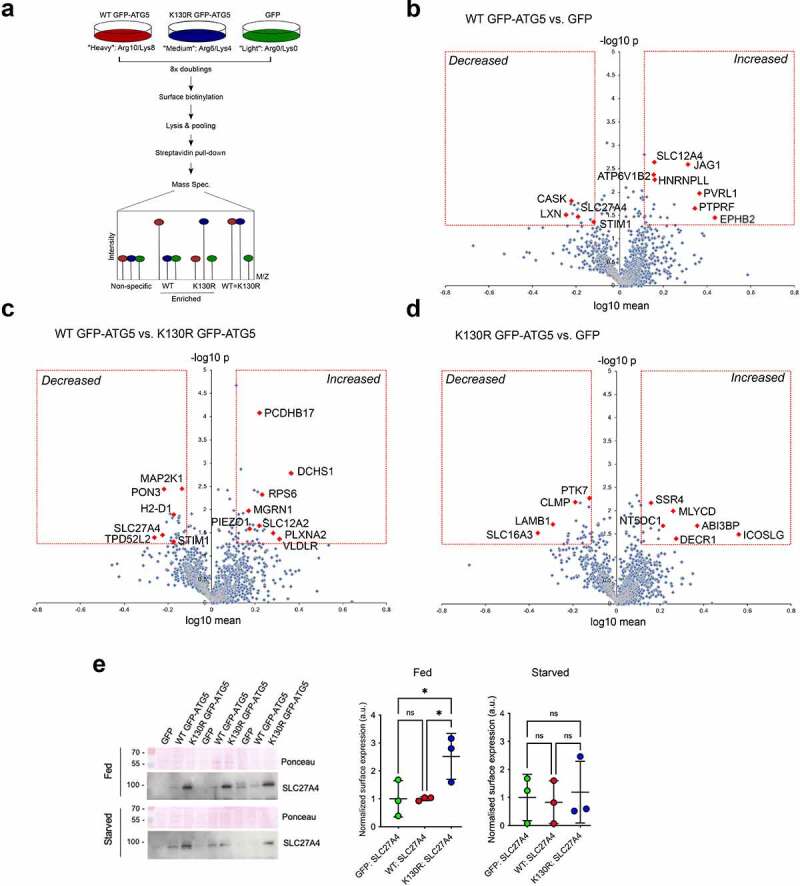
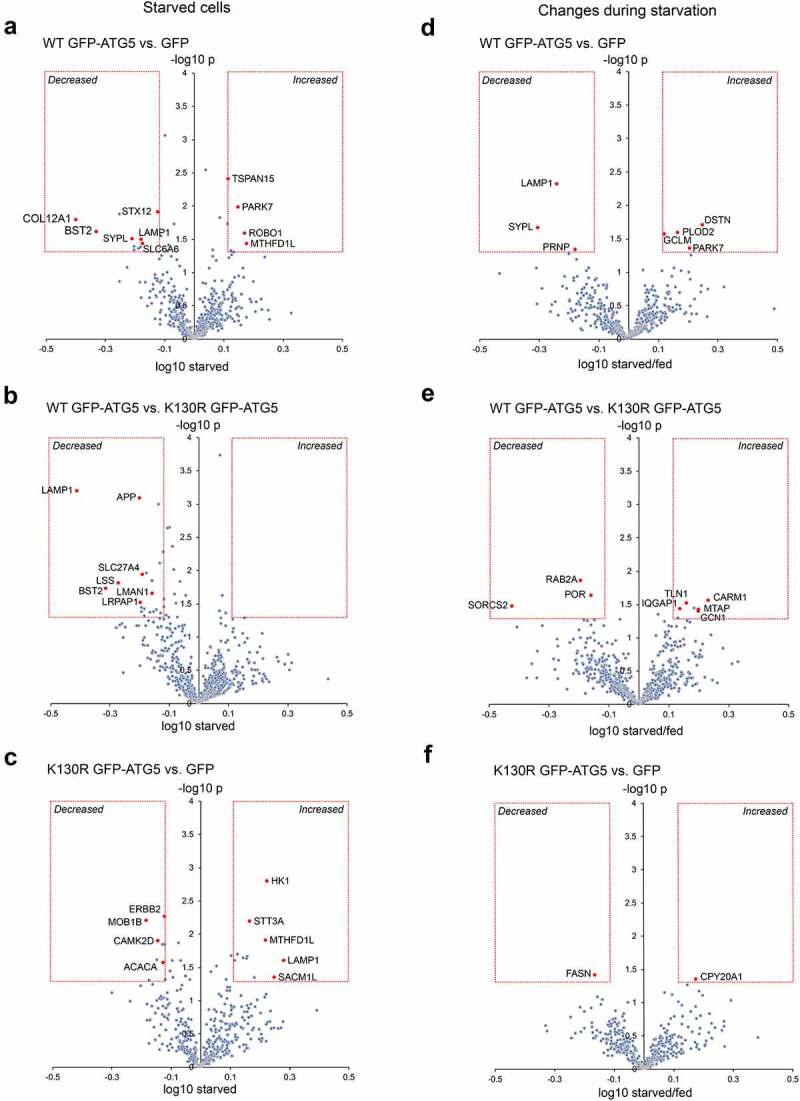
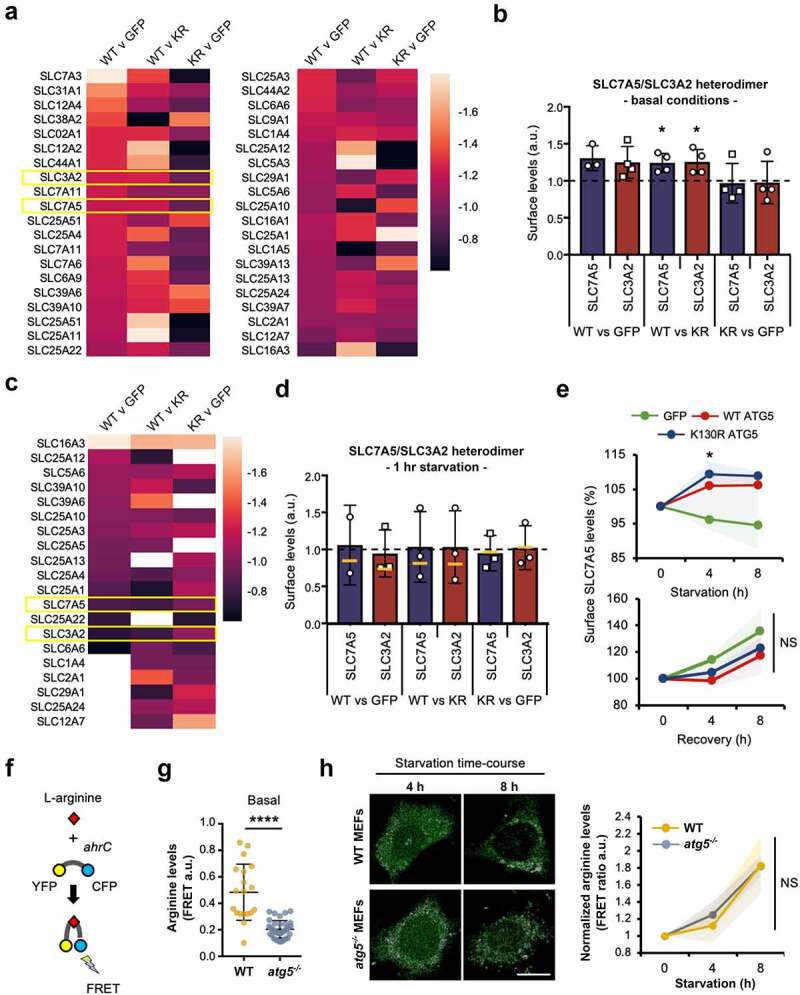
Autophagosome formation involves the sequential actions of conserved ATG proteins to coordinate the lipidation of the ubiquitin-like modifier Atg8-family proteins at the nascent phagophore membrane. Although the molecular steps driving this process are well understood, the source of membranes for the expanding phagophore and their mode of delivery are only now beginning to be revealed. Here, we have used quantitative SILAC-based proteomics to identify proteins that associate with the ATG12-ATG5 conjugate, a crucial player during Atg8-family protein lipidation. Our datasets reveal a strong enrichment of regulators of clathrin-mediated vesicular trafficking, including clathrin heavy and light chains, and several clathrin adaptors. Also identified were PIK3C2A (a phosphoinositide 3-kinase involved in clathrin-mediated endocytosis) and HIP1R (a component of clathrin vesicles), and the absence of either of these proteins alters autophagic flux in cell-based starvation assays. To determine whether the ATG12-ATG5 conjugate reciprocally influences trafficking within the endocytic compartment, we captured the cell surface proteomes of autophagy-competent and autophagy-incompetent mouse embryonic fibroblasts under fed and starved conditions. We report changes in the relative proportions of individual cell surface proteins and show that cell surface levels of the SLC7A5-SLC3A2 amino acid transporter are influenced by autophagy capability. Our data provide evidence for direct regulatory coupling between the ATG12-ATG5 conjugate and the clathrin membrane trafficking system and suggest candidate membrane proteins whose trafficking within the cell may be modulated by the autophagy machinery. Abbreviations: ATG, autophagy related; BafA1, bafilomycin A1; GFP, green fluorescent protein; HIP1R, huntingtin interacting protein 1 related; MEF, mouse embryo fibroblast; PIK3C2A, phosphatidylinositol-4-phosphate 3-kinase catalytic subunit type 2 alpha; SILAC, stable isotope labelling with amino acids in culture; SQSTM1, sequestosome 1; STRING, search tool for the retrieval of interacting genes/proteins.
Keywords: ATG12; ATG5; HIP1R; PIK3C2A; autophagy; clathrin; endocytosis; proteomics.
© 2022 The Author(s). Published by Informa UK Limited, trading as Taylor & Francis Group.
Conflict of interest statement
The authors report no potential competing interests.
Figures


Similar articles
-
RAB33B recruits the ATG16L1 complex to the phagophore via a noncanonical RAB binding protein.Autophagy. 2021 Sep;17(9):2290-2304. doi: 10.1080/15548627.2020.1822629. Epub 2020 Sep 22. Autophagy. 2021. PMID: 32960676 Free PMC article.
-
BIRC5/Survivin is a novel ATG12-ATG5 conjugate interactor and an autophagy-induced DNA damage suppressor in human cancer and mouse embryonic fibroblast cells.Autophagy. 2020 Jul;16(7):1296-1313. doi: 10.1080/15548627.2019.1671643. Epub 2019 Oct 15. Autophagy. 2020. PMID: 31612776 Free PMC article.
-
Dissecting the role of the Atg12-Atg5-Atg16 complex during autophagosome formation.Autophagy. 2013 Mar;9(3):424-5. doi: 10.4161/auto.22931. Epub 2013 Jan 15. Autophagy. 2013. PMID: 23321721 Free PMC article.
-
Two ubiquitin-like conjugation systems that mediate membrane formation during autophagy.Essays Biochem. 2013;55:39-50. doi: 10.1042/bse0550039. Essays Biochem. 2013. PMID: 24070470 Review.
-
Autophagy in the physiological endometrium and cancer.Autophagy. 2021 May;17(5):1077-1095. doi: 10.1080/15548627.2020.1752548. Epub 2020 May 13. Autophagy. 2021. PMID: 32401642 Free PMC article. Review.
Cited by
-
Noncanonical roles of ATG5 and membrane atg8ylation in retromer assembly and function.Elife. 2025 Jan 7;13:RP100928. doi: 10.7554/eLife.100928. Elife. 2025. PMID: 39773872 Free PMC article.
-
Atg8-Family Proteins-Structural Features and Molecular Interactions in Autophagy and Beyond.Cells. 2020 Sep 1;9(9):2008. doi: 10.3390/cells9092008. Cells. 2020. PMID: 32882854 Free PMC article. Review.
-
Lack of GABARAP-Type Proteins Is Accompanied by Altered Golgi Morphology and Surfaceome Composition.Int J Mol Sci. 2020 Dec 23;22(1):85. doi: 10.3390/ijms22010085. Int J Mol Sci. 2020. PMID: 33374830 Free PMC article.
-
ATG5 selectively engages virus-tethered BST2/tetherin in an LC3C-associated pathway.Proc Natl Acad Sci U S A. 2023 May 16;120(20):e2217451120. doi: 10.1073/pnas.2217451120. Epub 2023 May 8. Proc Natl Acad Sci U S A. 2023. PMID: 37155854 Free PMC article.
-
Noncanonical roles of ATG5 and membrane atg8ylation in retromer assembly and function.bioRxiv [Preprint]. 2024 Oct 14:2024.07.10.602886. doi: 10.1101/2024.07.10.602886. bioRxiv. 2024. Update in: Elife. 2025 Jan 07;13:RP100928. doi: 10.7554/eLife.100928. PMID: 39026874 Free PMC article. Updated. Preprint.
References
-
- Dikic, I. and Elazar Z., Mechanism and medical implications of mammalian autophagy. Nat Rev Mol Cell Biol, 2018. 19(6): p. 349–364. - PubMed
-
- Nakatogawa, H., Two ubiquitin-like conjugation systems that mediate membrane formation during autophagy. Essays Biochem, 2013. 55: p. 39–50. - PubMed
-
- Lamb, C.A., Yoshimori T., and Tooze S.A., The autophagosome: origins unknown, biogenesis complex. Nat Rev Mol Cell Biol, 2013. 14(12): p. 759–74. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials