

HMCES protects immunoglobulin genes specifically from deletions during somatic hypermutation
- PMID: 35450882
- PMCID: PMC9067407
- DOI: 10.1101/gad.349438.122
HMCES protects immunoglobulin genes specifically from deletions during somatic hypermutation
Abstract
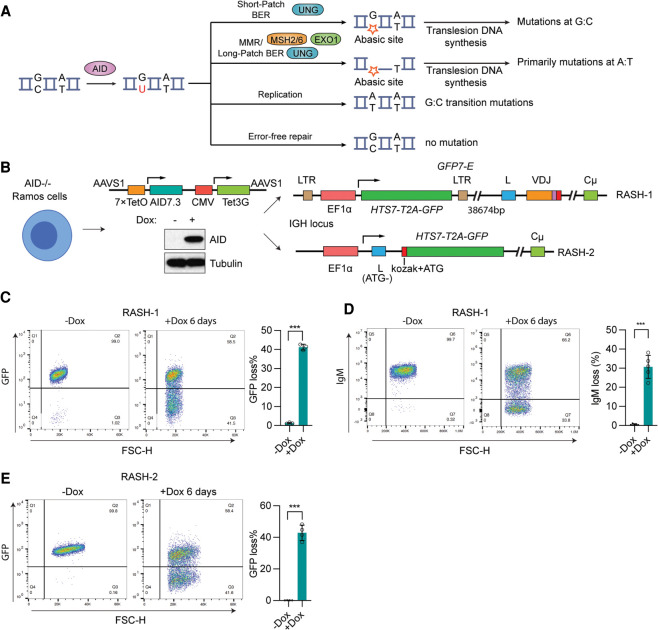
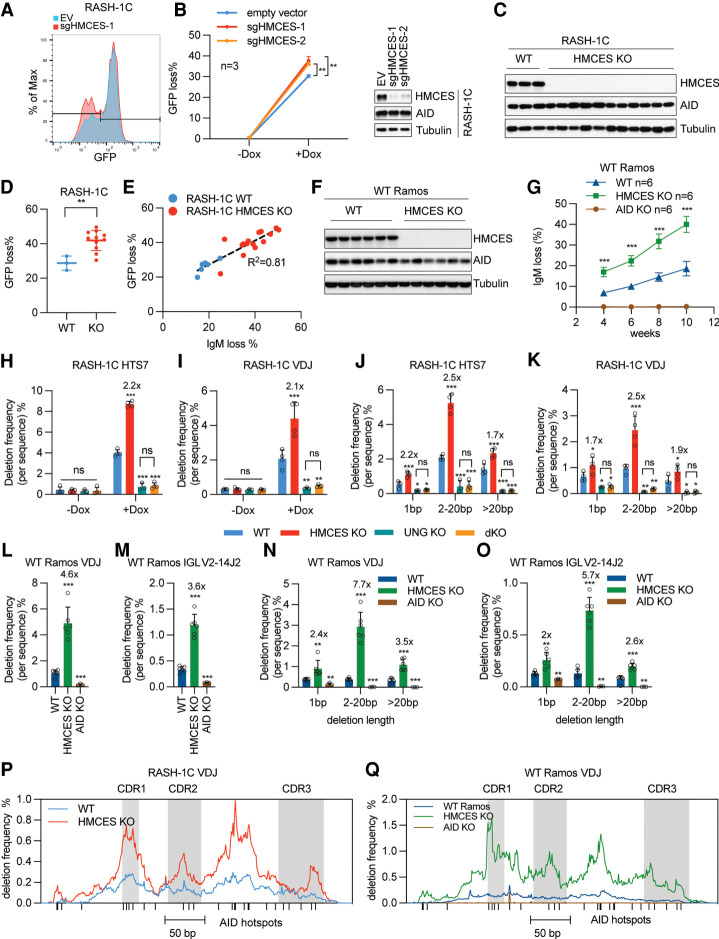
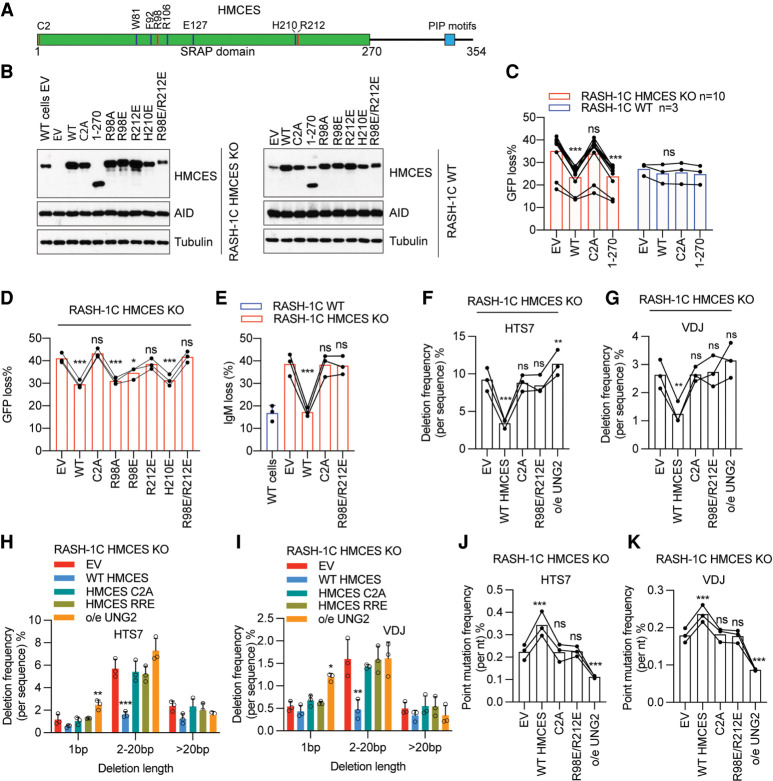
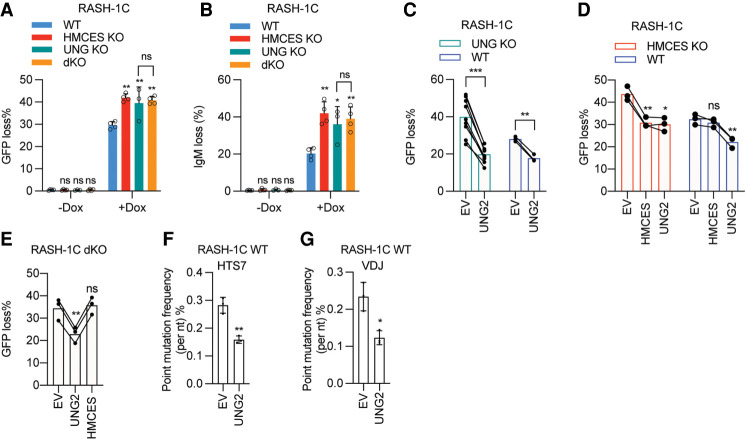
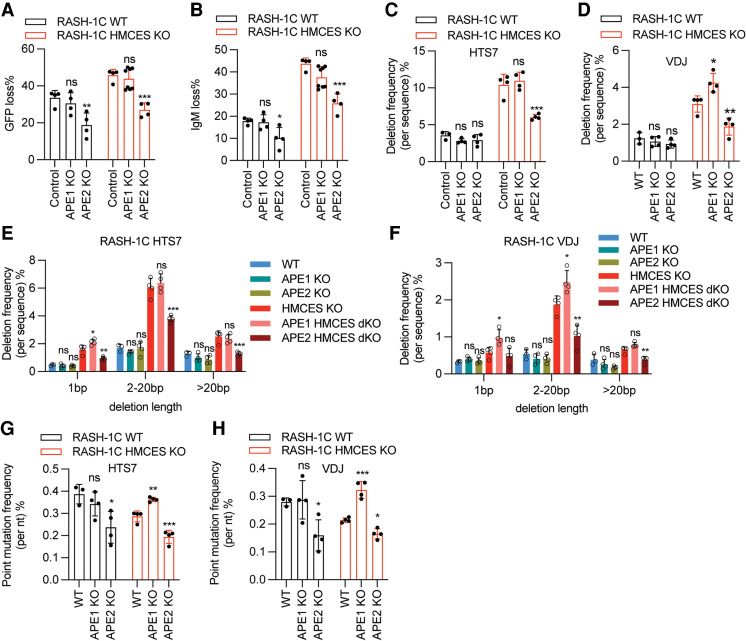
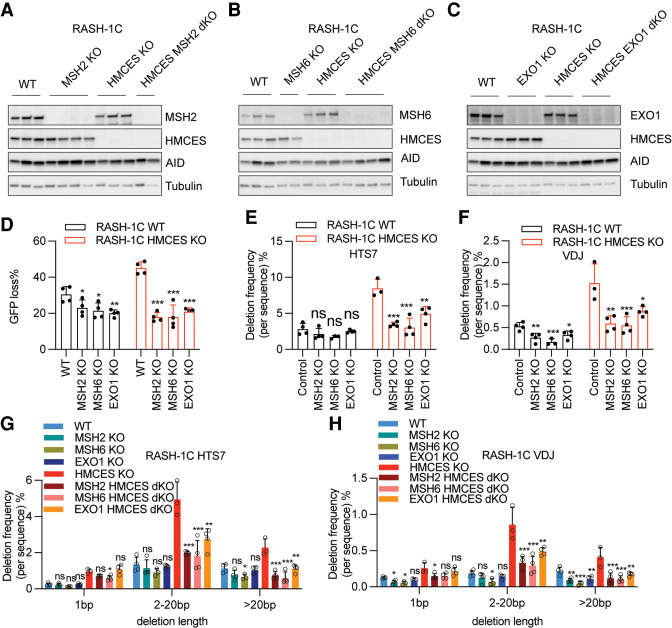
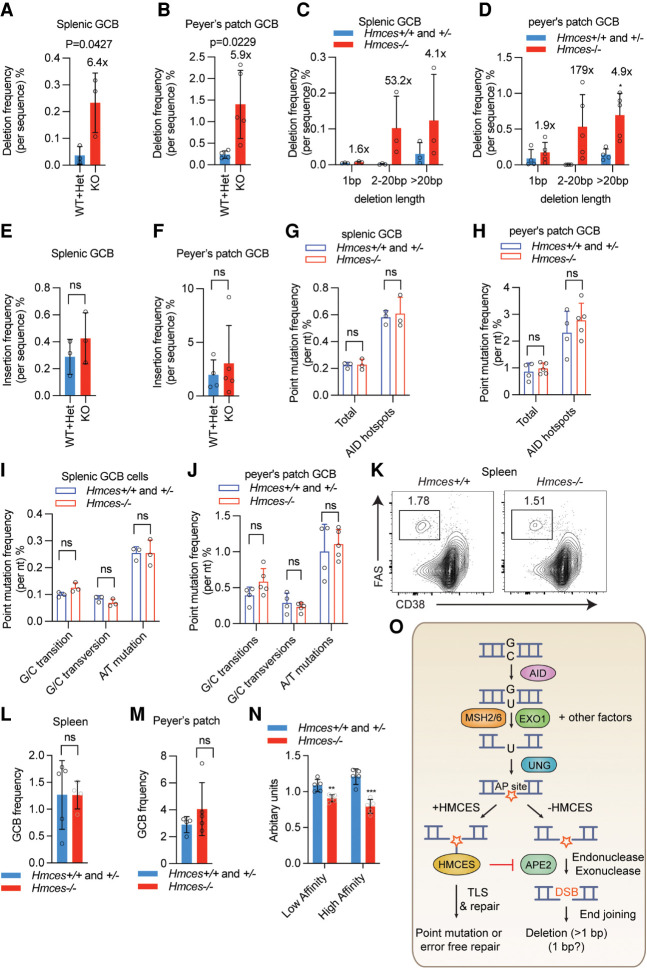
Somatic hypermutation (SHM) produces point mutations in immunoglobulin (Ig) genes in B cells when uracils created by the activation-induced deaminase are processed in a mutagenic manner by enzymes of the base excision repair (BER) and mismatch repair (MMR) pathways. Such uracil processing creates DNA strand breaks and is susceptible to the generation of deleterious deletions. Here, we demonstrate that the DNA repair factor HMCES strongly suppresses deletions without significantly affecting other parameters of SHM in mouse and human B cells, thereby facilitating the production of antigen-specific antibodies. The deletion-prone repair pathway suppressed by HMCES operates downstream from the uracil glycosylase UNG and is mediated by the combined action of BER factor APE2 and MMR factors MSH2, MSH6, and EXO1. HMCES's ability to shield against deletions during SHM requires its capacity to form covalent cross-links with abasic sites, in sharp contrast to its DNA end-joining role in class switch recombination but analogous to its genome-stabilizing role during DNA replication. Our findings lead to a novel model for the protection of Ig gene integrity during SHM in which abasic site cross-linking by HMCES intercedes at a critical juncture during processing of vulnerable gapped DNA intermediates by BER and MMR enzymes.
Keywords: AID; HMCES; antibody affinity maturation; base excision repair; mismatch repair; somatic hypermutation.
© 2022 Wu et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous