

αα-hub coregulator structure and flexibility determine transcription factor binding and selection in regulatory interactomes
- PMID: 35452682
- PMCID: PMC9127584
- DOI: 10.1016/j.jbc.2022.101963
αα-hub coregulator structure and flexibility determine transcription factor binding and selection in regulatory interactomes
Abstract
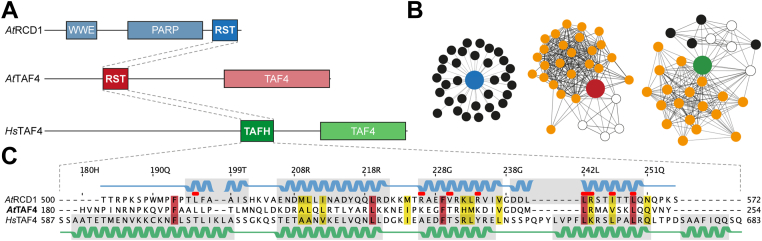
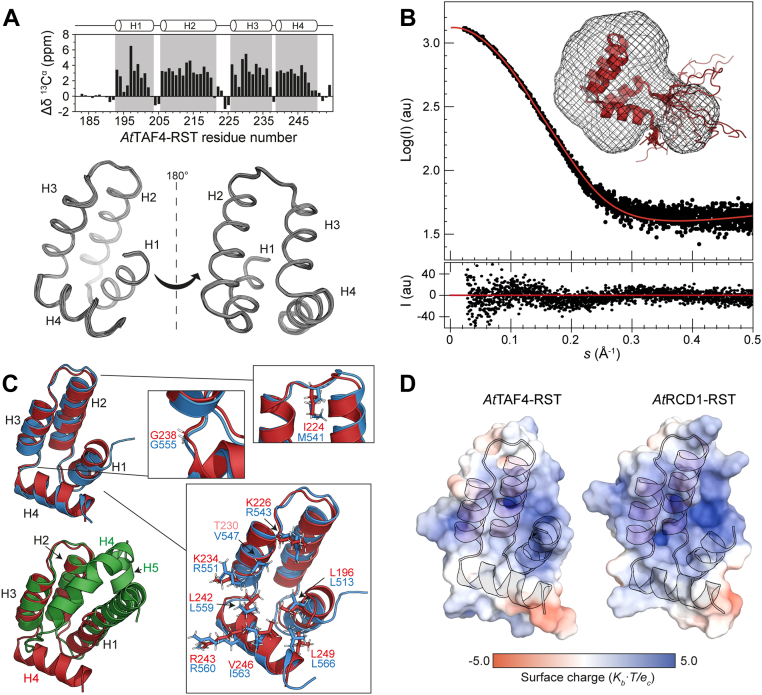
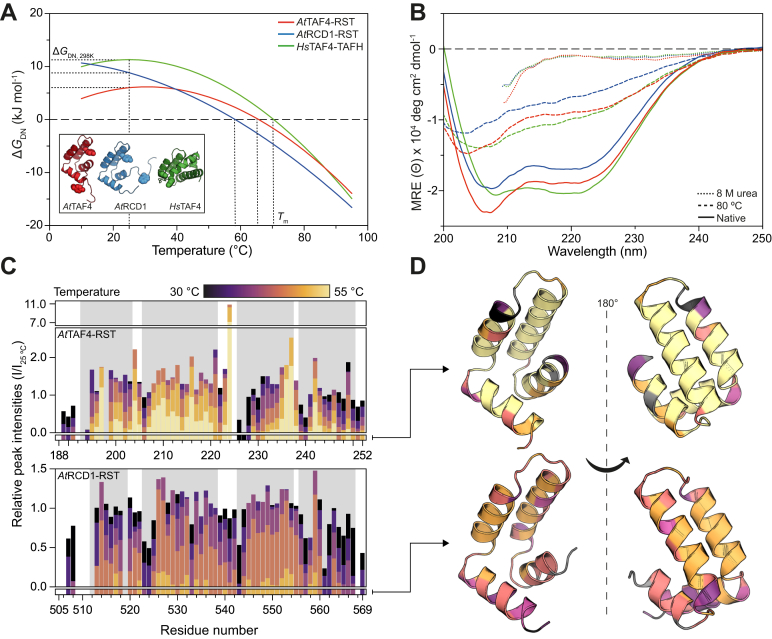
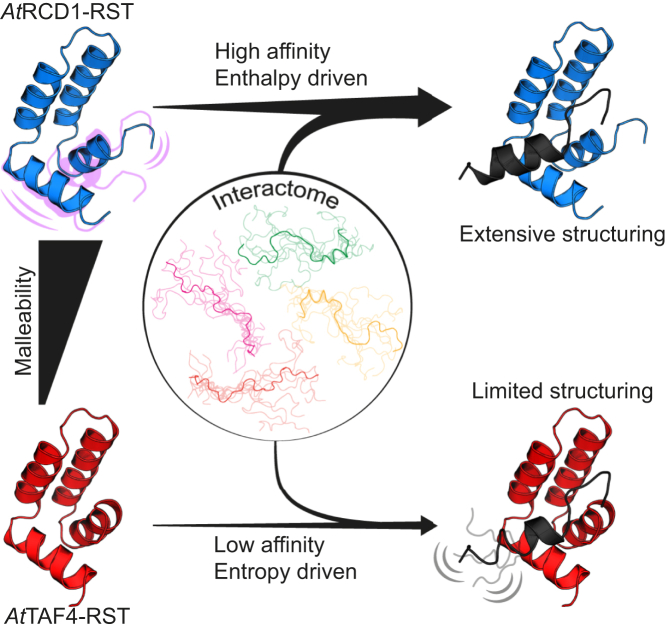
Formation of transcription factor (TF)-coregulator complexes is a key step in transcriptional regulation, with coregulators having essential functions as hub nodes in molecular networks. How specificity and selectivity are maintained in these nodes remain open questions. In this work, we addressed specificity in transcriptional networks using complexes formed between TFs and αα-hubs, which are defined by a common αα-hairpin secondary structure motif, as a model. Using NMR spectroscopy and binding thermodynamics, we analyzed the structure, dynamics, stability, and ligand-binding properties of the Arabidopsis thaliana RST domains from TAF4 and known binding partner RCD1, and the TAFH domain from human TAF4, allowing comparison across species, functions, and architectural contexts. While these αα-hubs shared the αα-hairpin motif, they differed in length and orientation of accessory helices as well as in their thermodynamic profiles of ligand binding. Whereas biologically relevant RCD1-ligand pairs displayed high affinity driven by enthalpy, TAF4-ligand interactions were entropy driven and exhibited less binding-induced structuring. We in addition identified a thermal unfolding state with a structured core for all three domains, although the temperature sensitivity differed. Thermal stability studies suggested that initial unfolding of the RCD1-RST domain localized around helix 1, lending this region structural malleability, while effects in TAF4-RST were more stochastic, suggesting variability in structural adaptability upon binding. Collectively, our results support a model in which hub structure, flexibility, and binding thermodynamics contribute to αα-hub-TF binding specificity, a finding of general relevance to the understanding of coregulator-ligand interactions and interactome sizes.
Keywords: alternative state; entropy; interactome size; intrinsically disordered protein; malleability; protein dynamic; protein structure; protein–protein interaction; transcriptional coactivator; αα-hub specificity.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures


References
-
- Ma J., Ptashne M. A new class of yeast transcriptional activators. Cell. 1987;51:113–119. - PubMed
-
- Kundu T.K., Palhan V.B., Wang Z., An W., Cole P.A., Roeder R.G. Activator-dependent transcription from chromatin in vitro involving targeted histone acetylation by p300. Mol. Cell. 2000;6:551–561. - PubMed
-
- Ptashne M., Gann A. Transcriptional activation by recruitment. Nature. 1997;386:569–577. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
