

Lack of VMP1 impairs hepatic lipoprotein secretion and promotes non-alcoholic steatohepatitis
- PMID: 35452693
- PMCID: PMC9449865
- DOI: 10.1016/j.jhep.2022.04.010
Lack of VMP1 impairs hepatic lipoprotein secretion and promotes non-alcoholic steatohepatitis
Abstract
Background & aims: Vacuole membrane protein 1 (VMP1) is an endoplasmic reticulum (ER) transmembrane protein that regulates the formation of autophagosomes and lipid droplets. Recent evidence suggests that VMP1 plays a critical role in lipoprotein secretion in zebra fish and cultured cells. However, the pathophysiological roles and mechanisms by which VMP1 regulates lipoprotein secretion and lipid accumulation in non-alcoholic fatty liver disease (NAFLD) and non-alcoholic steatohepatitis (NASH) are unknown.
Methods: Liver-specific and hepatocyte-specific Vmp1 knockout mice as well as Vmp1 knock-in mice were generated by crossing Vmp1flox or Vmp1KI mice with albumin-Cre mice or by injecting AAV8-TBG-cre, respectively. Lipid and energy metabolism in these mice were characterized by metabolomic and transcriptome analyses. Mice with hepatic overexpression of VMP1 who were fed a NASH diet were also characterized.
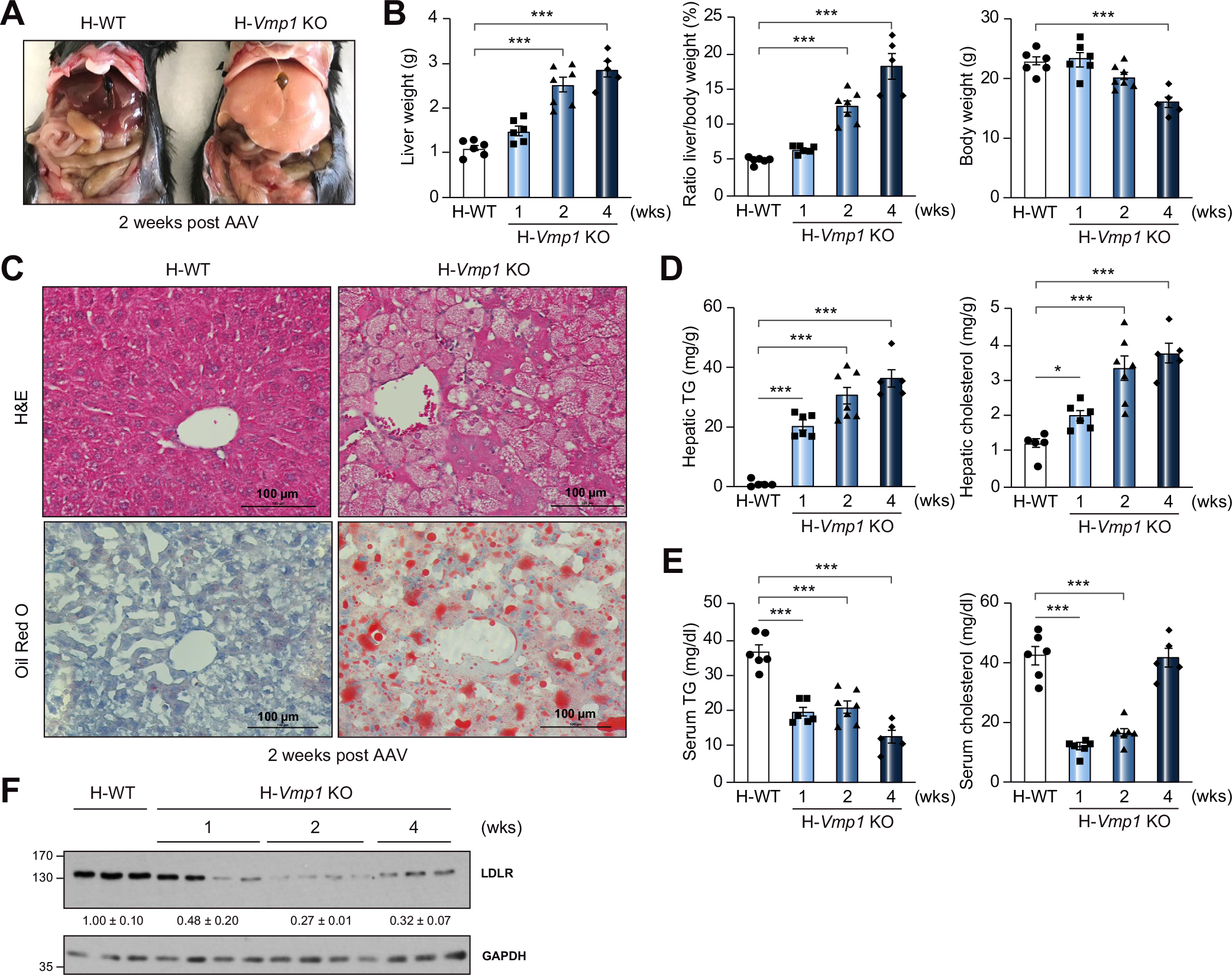
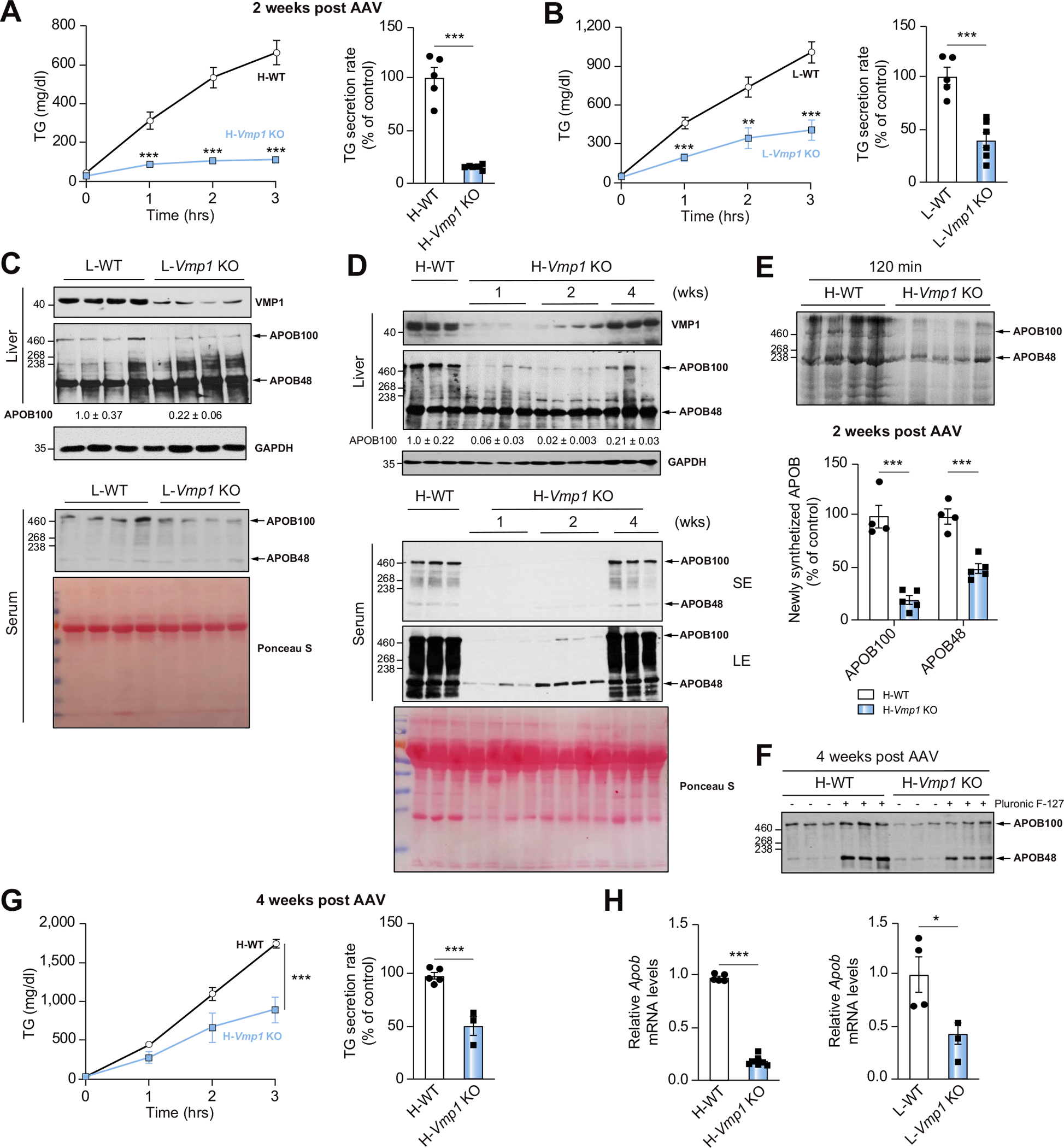
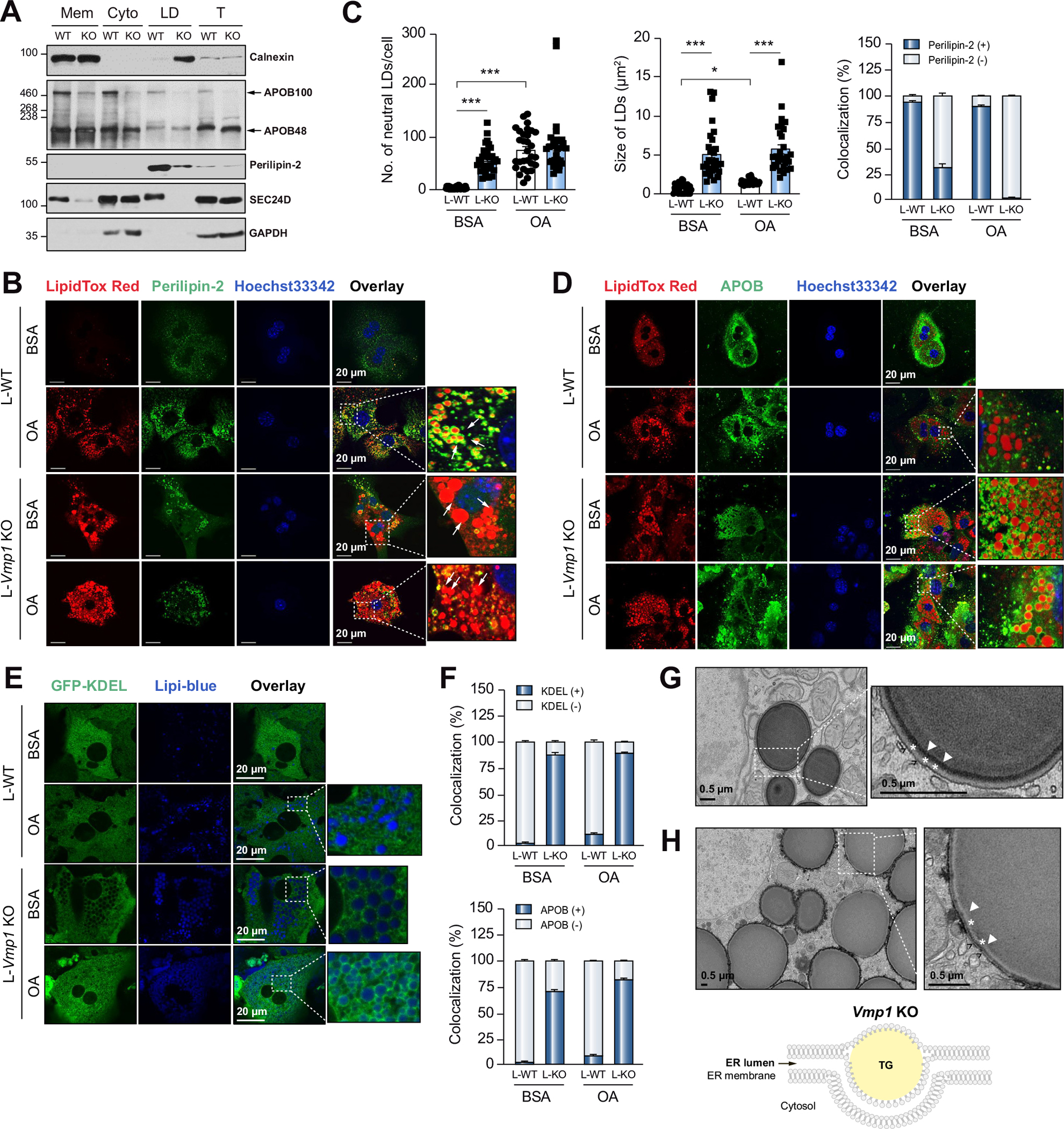
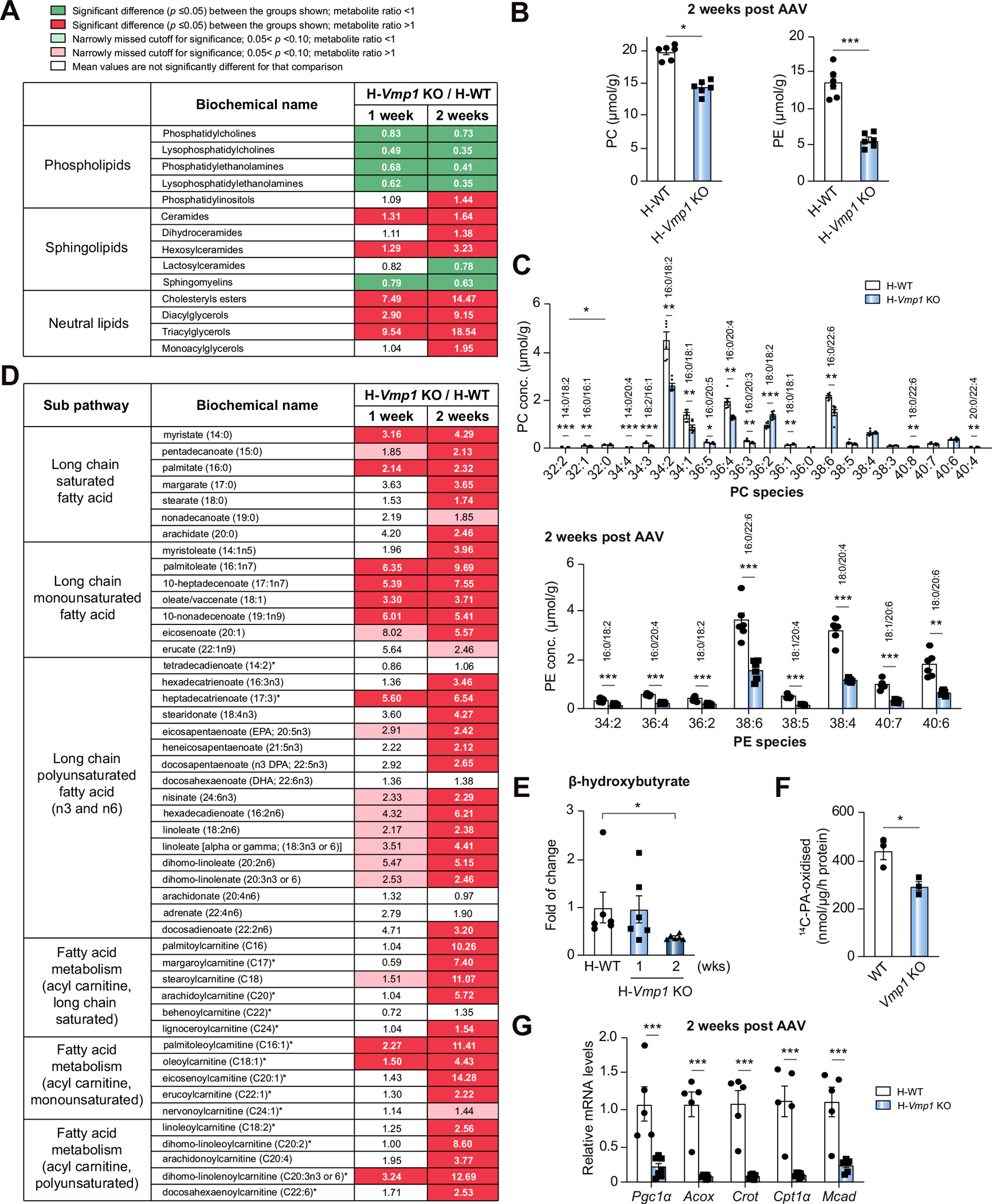
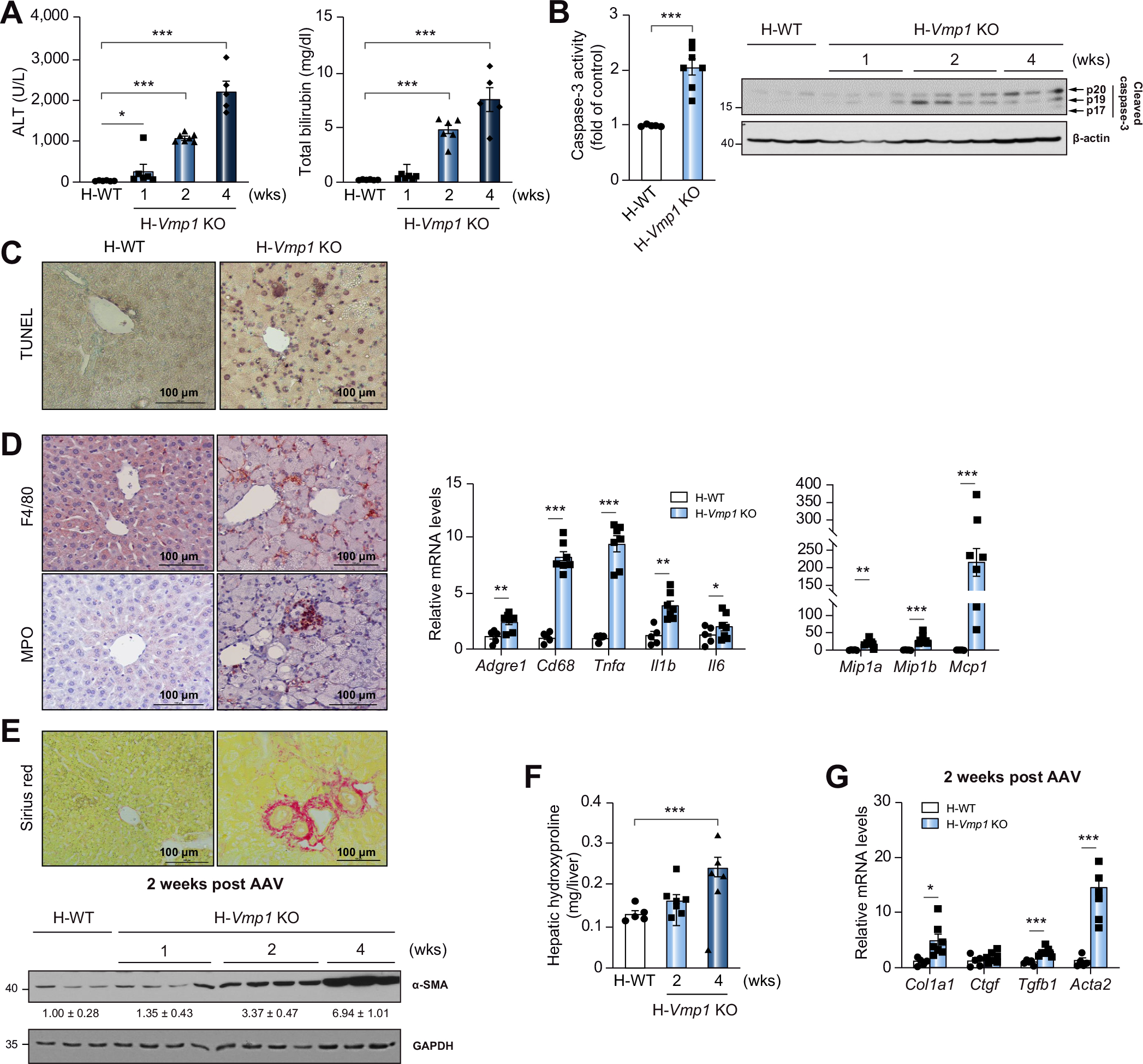
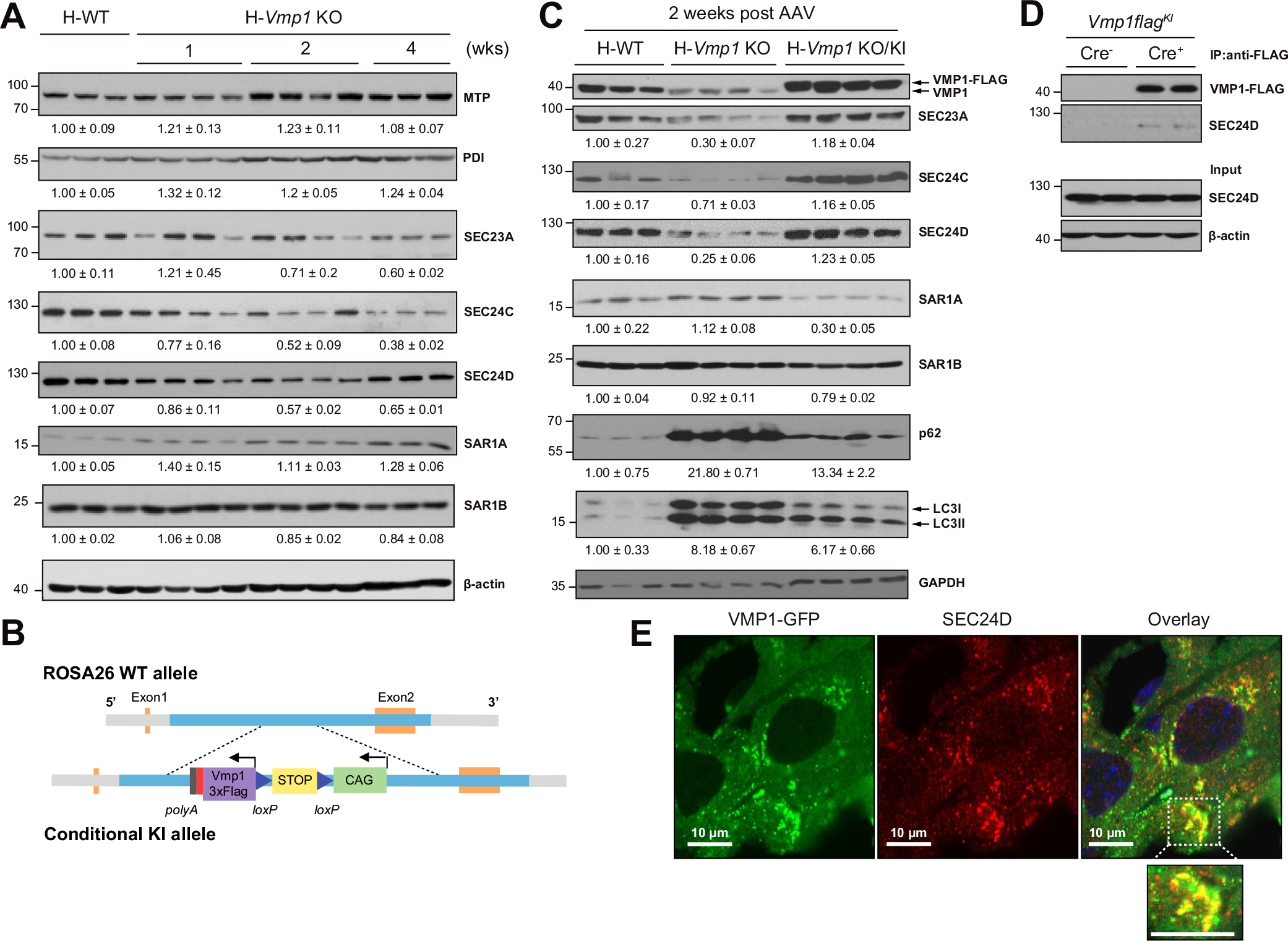
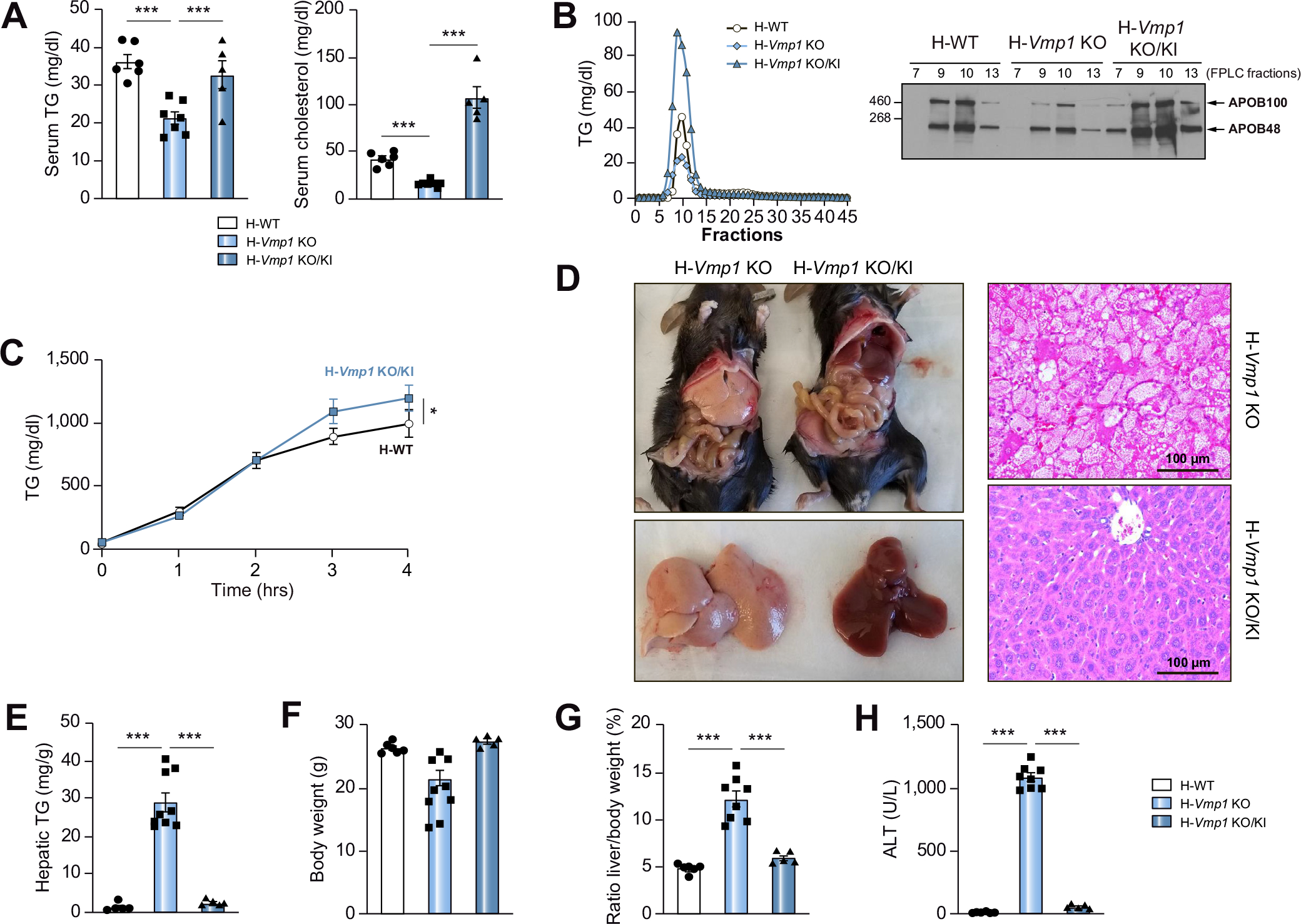
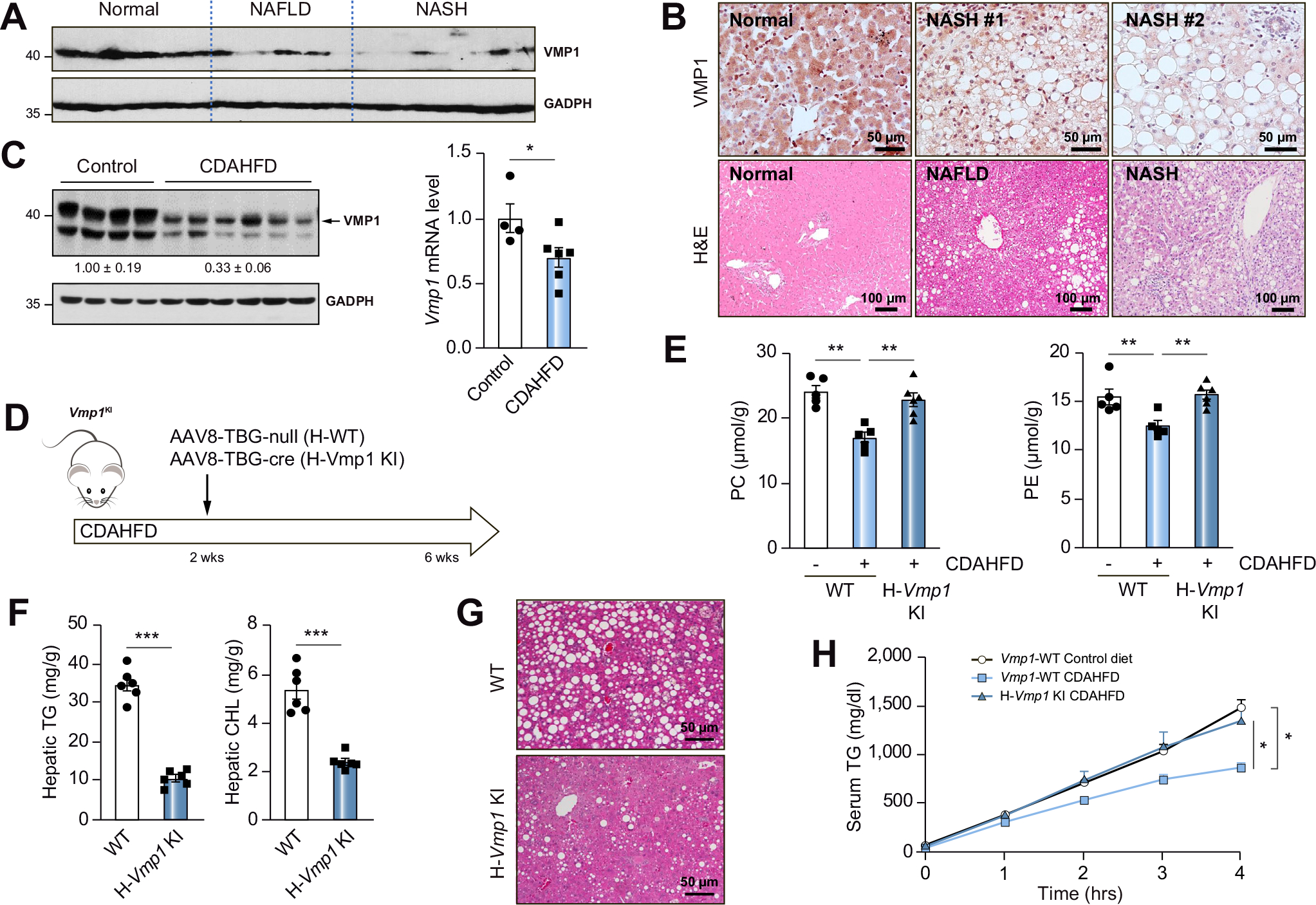
Results: Hepatocyte-specific deletion of Vmp1 severely impaired VLDL secretion resulting in massive hepatic steatosis, hepatocyte death, inflammation and fibrosis, which are hallmarks of NASH. Mechanistically, loss of Vmp1 led to decreased hepatic levels of phosphatidylcholine and phosphatidylethanolamine as well as to changes in phospholipid composition. Deletion of Vmp1 in mouse liver also led to the accumulation of neutral lipids in the ER bilayer and impaired mitochondrial beta-oxidation. Overexpression of VMP1 ameliorated steatosis in diet-induced NASH by improving VLDL secretion. Importantly, we also showed that decreased liver VMP1 is associated with NAFLD/NASH in humans.
Conclusions: Our results provide novel insights on the role of VMP1 in regulating hepatic phospholipid synthesis and lipoprotein secretion in the pathogenesis of NAFLD/NASH.
Lay summary: Non-alcoholic fatty liver disease and its more severe form, non-alcoholic steatohepatitis, are associated with a build-up of fat in the liver (steatosis). However, the exact mechanisms that underly steatosis in patients are not completely understood. Herein, the authors identified that the lack of a protein called VMP1 impairs the secretion and metabolism of fats in the liver and could therefore contribute to the development and progression of non-alcoholic fatty liver disease.
Keywords: NAFLD; VLDL; autophagy; endoplasmic reticulum; liver injury.
Copyright © 2022 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Conflict of interest The authors who have taken part in this study declare that they have nothing to disclose. Please refer to the accompanying ICMJE disclosure forms for further details.
Figures
References
-
- Davidson NO, Shelness GS. APOLIPOPROTEIN B: mRNA editing, lipoprotein assembly, and presecretory degradation. Annu Rev Nutr 2000;20:169–193. - PubMed
-
- Dusetti NJ, Jiang Y, Vaccaro MI, Tomasini R, Azizi Samir A, Calvo EL, et al. Cloning and expression of the rat vacuole membrane protein 1 (VMP1), a new gene activated in pancreas with acute pancreatitis, which promotes vacuole formation. Biochem Biophys Res Commun 2002;290:641–649. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R21 AA026904/AA/NIAAA NIH HHS/United States
- R01 DK106540/DK/NIDDK NIH HHS/United States
- R56 DK129234/DK/NIDDK NIH HHS/United States
- K01 DK112967/DK/NIDDK NIH HHS/United States
- R01 DK124612/DK/NIDDK NIH HHS/United States
- R01 AA020518/AA/NIAAA NIH HHS/United States
- R37 AA020518/AA/NIAAA NIH HHS/United States
- U54 HD090216/HD/NICHD NIH HHS/United States
- R01 DK131064/DK/NIDDK NIH HHS/United States
- R01 DK117965/DK/NIDDK NIH HHS/United States
- U01 AA024733/AA/NIAAA NIH HHS/United States
- P01 HL151328/HL/NHLBI NIH HHS/United States
- P30 GM118247/GM/NIGMS NIH HHS/United States
- R01 DK119437/DK/NIDDK NIH HHS/United States
- P20 GM144269/GM/NIGMS NIH HHS/United States
- P30 DK020579/DK/NIDDK NIH HHS/United States
- R01 DK121970/DK/NIDDK NIH HHS/United States
- P30 DK052574/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
