

Respiratory Burst Oxidase Homolog D as a Modulating Component of Oxidative Response under Ammonium Toxicity
- PMID: 35453389
- PMCID: PMC9031508
- DOI: 10.3390/antiox11040703
Respiratory Burst Oxidase Homolog D as a Modulating Component of Oxidative Response under Ammonium Toxicity
Abstract
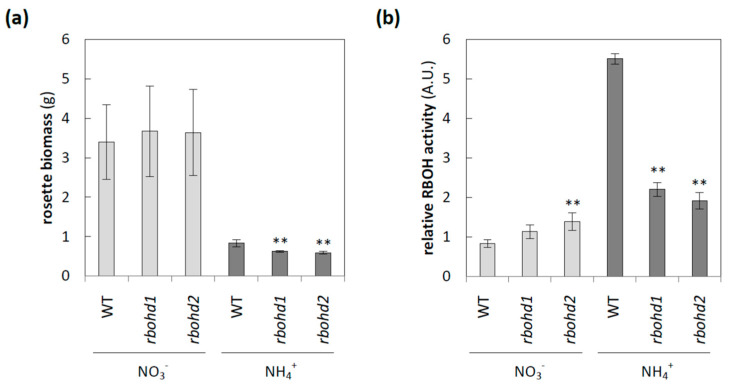
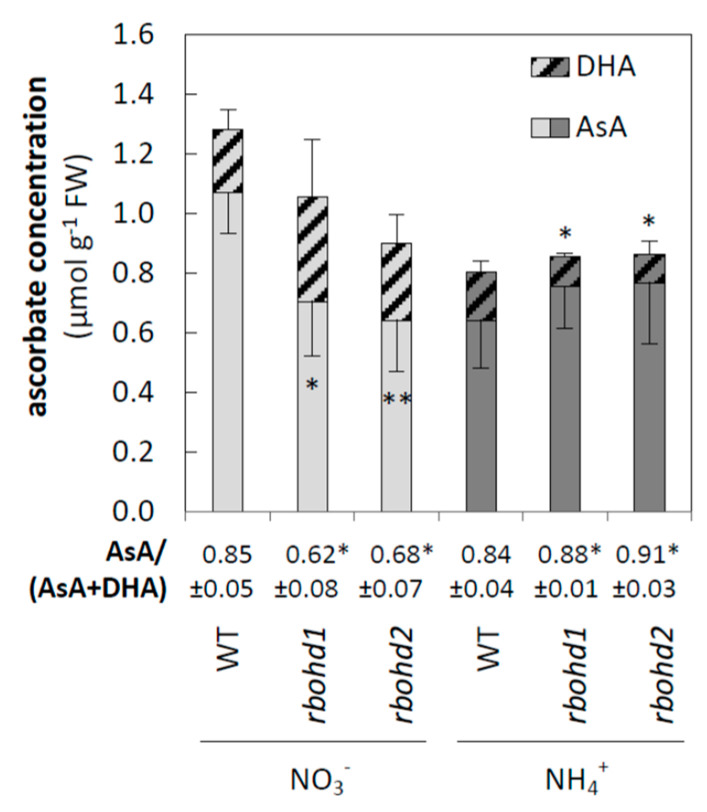
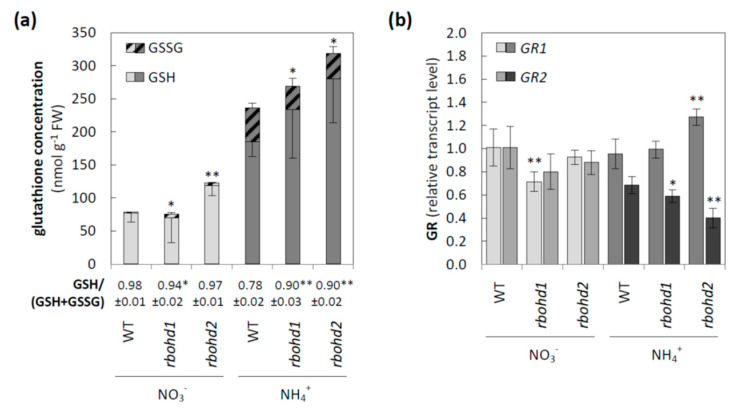
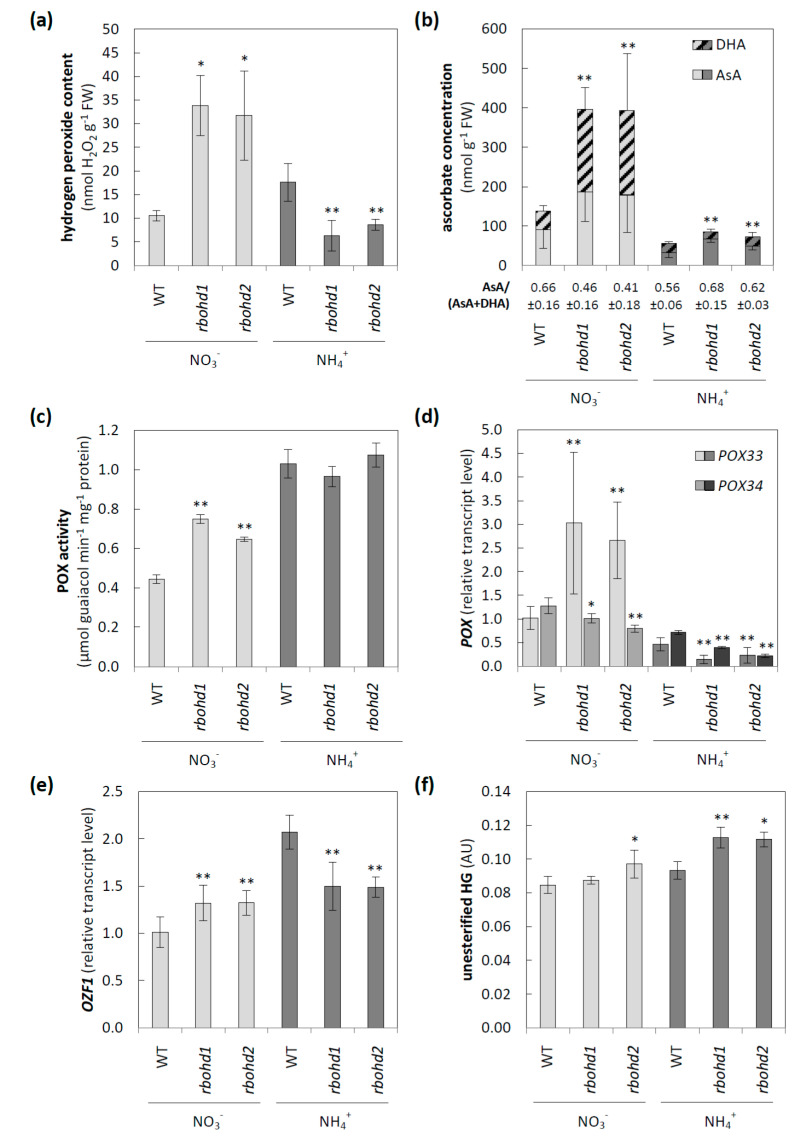
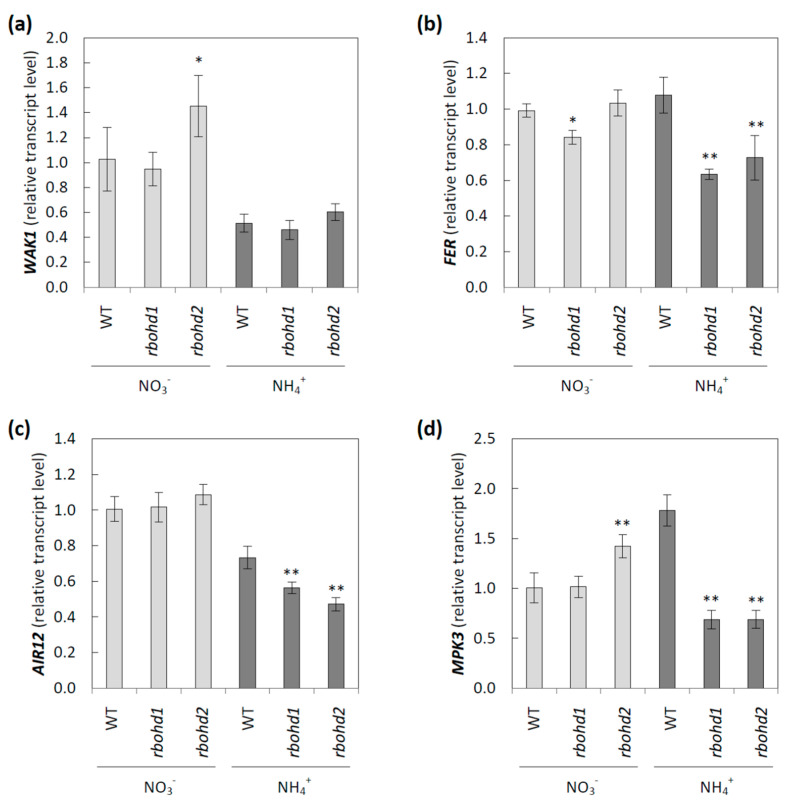
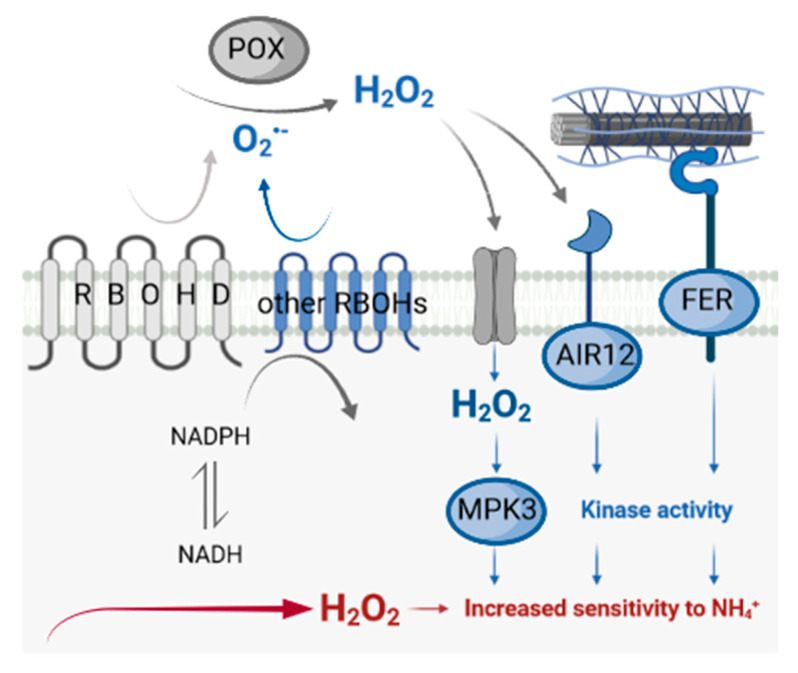
Delayed growth, a visible phenotypic component of the so-called ammonium syndrome, occurs when ammonium is the sole inorganic nitrogen source. Previously, we have shown that modification of apoplastic reactive oxygen species (apROS) metabolism is a key factor contributing to plant growth retardation under ammonium nutrition. Here, we further analyzed the changes in apROS metabolism in transgenic plants with disruption of the D isoform of the respiratory burst oxidase homolog (RBOH) that is responsible for apROS production. Ammonium-grown Arabidopsisrbohd plants are characterized by up to 50% lower contents of apoplastic superoxide and hydrogen peroxide. apROS sensing markers such as OZF1 and AIR12 were downregulated, and the ROS-responsive signaling pathway, including MPK3, was also downregulated in rbohd plants cultivated using ammonium as the sole nitrogen source. Additionally, the expression of the cell-wall-integrity marker FER and peroxidases 33 and 34 was decreased. These modifications may contribute to phenomenon wherein ammonium inhibited the growth of transgenic plants to a greater extent than that of wild-type plants. Overall, this study indicated that due to disruption of apROS metabolism, rbohd plants cannot adjust to ammonium toxicity and are more sensitive to these conditions.
Keywords: ammonium syndrome; apoplastic antioxidants; apoplastic reactive oxygen species metabolism; growth retardation; respiratory burst oxidase homolog.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Kangasjärvi S., Kangasjärvi J. Towards Understanding Extracellular ROS Sensory and Signaling Systems in Plants. Adv. Bot. 2014;2014:538946. doi: 10.1155/2014/538946. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
