Cellular Distribution of Brain Aquaporins and Their Contribution to Cerebrospinal Fluid Homeostasis and Hydrocephalus
- PMID: 35454119
- PMCID: PMC9025855
- DOI: 10.3390/biom12040530
Cellular Distribution of Brain Aquaporins and Their Contribution to Cerebrospinal Fluid Homeostasis and Hydrocephalus
Abstract
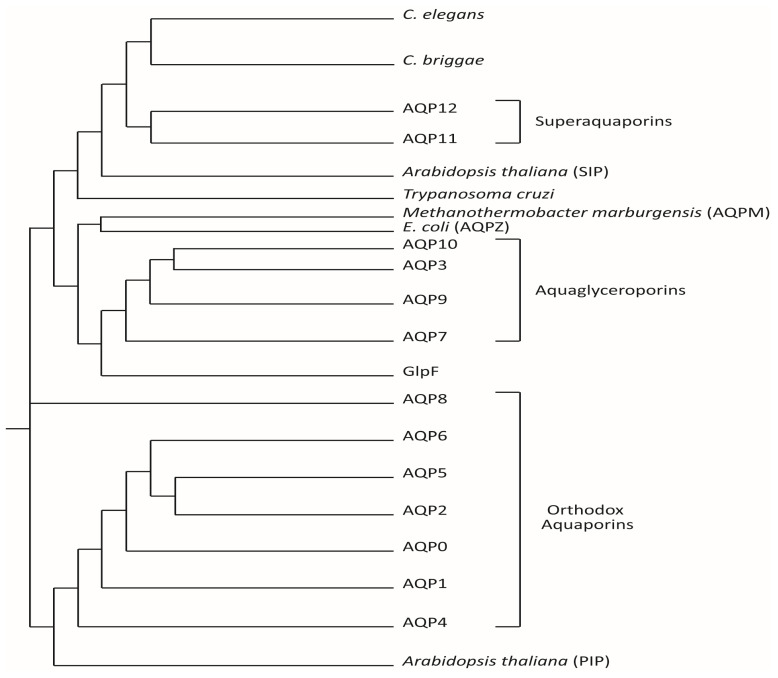
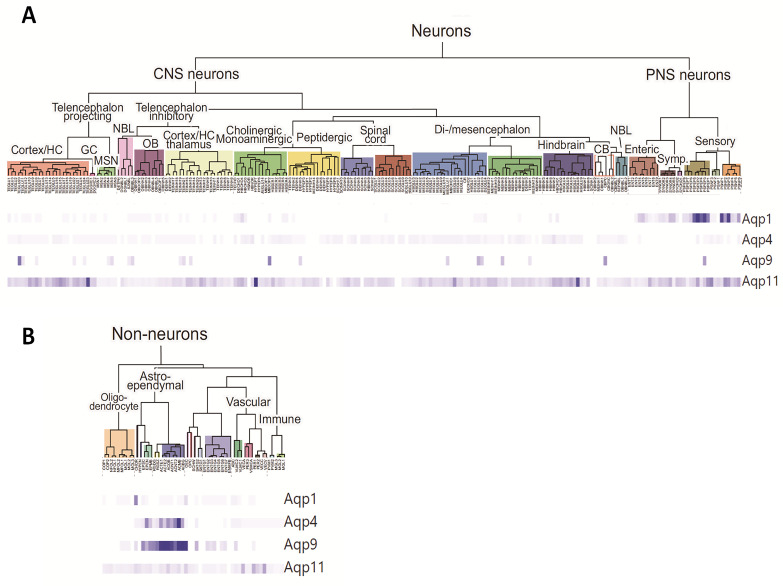
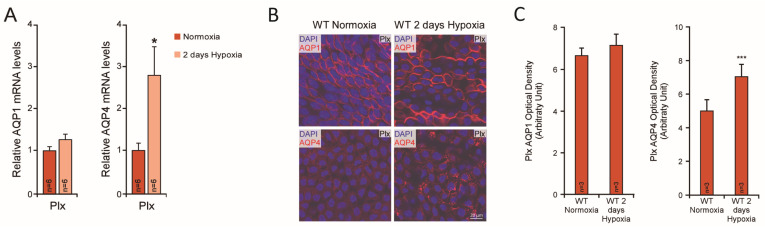
Brain aquaporins facilitate the movement of water between the four water compartments: blood, cerebrospinal fluid, interstitial fluid, and intracellular fluid. This work analyzes the expression of the four most abundant aquaporins (AQPs) (AQP1, AQP4, AQP9, and AQP11) in the brains of mice and discuss their contribution to hydrocephalus. We analyzed available data from single-cell RNA sequencing of the central nervous system of mice to describe the expression of aquaporins and compare their distribution with that based on qPCR, western blot, and immunohistochemistry assays. Expression of AQP1 in the apical cell membrane of choroid plexus epithelial cells and of AQP4 in ependymal cells, glia limitans, and astrocyte processes in the pericapillary end foot is consistent with the involvement of both proteins in cerebrospinal fluid homeostasis. The expression of both aquaporins compensates for experimentally induced hydrocephalus in the animals. Recent data demonstrate that hypoxia in aged animals alters AQP4 expression in the choroidal plexus and cortex, increasing the ventricle size and intraventricular pressure. Cerebral distensibility is reduced in parallel with a reduction in cerebrospinal fluid drainage and cognitive deterioration. We propose that aged mice chronically exposed to hypoxia represent an excellent experimental model for studying the pathophysiological characteristics of idiopathic normal pressure hydrocephalus and roles for AQPs in such disease.
Keywords: aging; aquaporins; cerebrospinal fluid; choroid plexus; ependyma; hydrocephalus; hypoxia; sequencing; single-cell RNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures