

Effects of a Short-Term Lipopolysaccharides Challenge on Mouse Brain and Liver Peroxisomal Antioxidant and β-oxidative Functions: Protective Action of Argan Oil
- PMID: 35455460
- PMCID: PMC9030085
- DOI: 10.3390/ph15040465
Effects of a Short-Term Lipopolysaccharides Challenge on Mouse Brain and Liver Peroxisomal Antioxidant and β-oxidative Functions: Protective Action of Argan Oil
Abstract
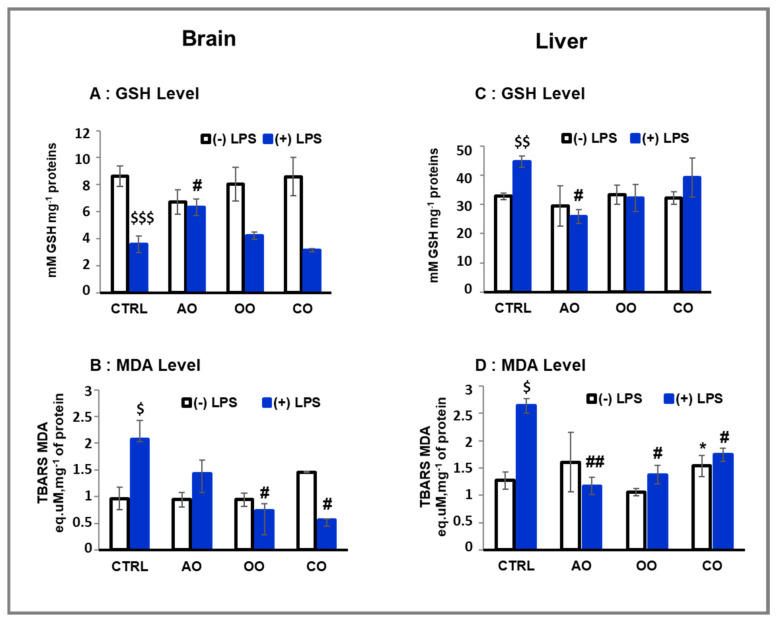
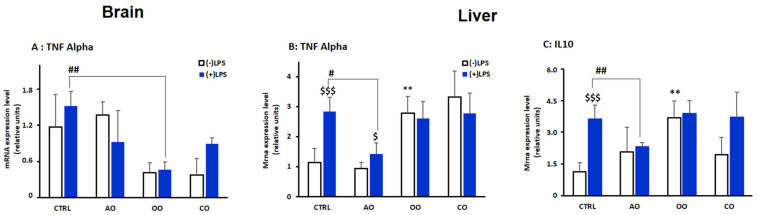
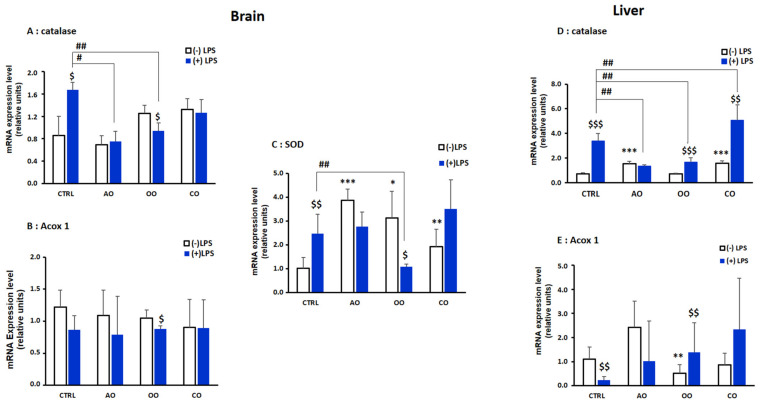
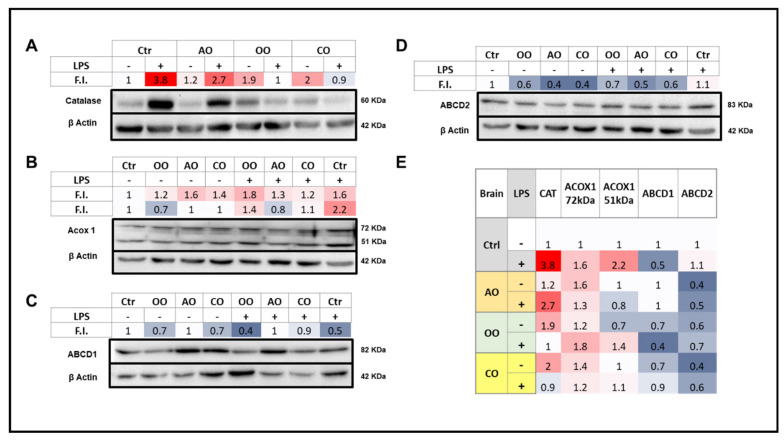
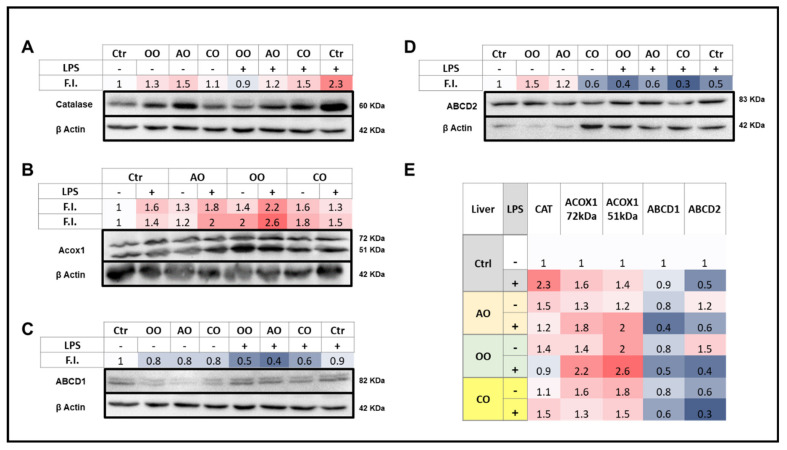
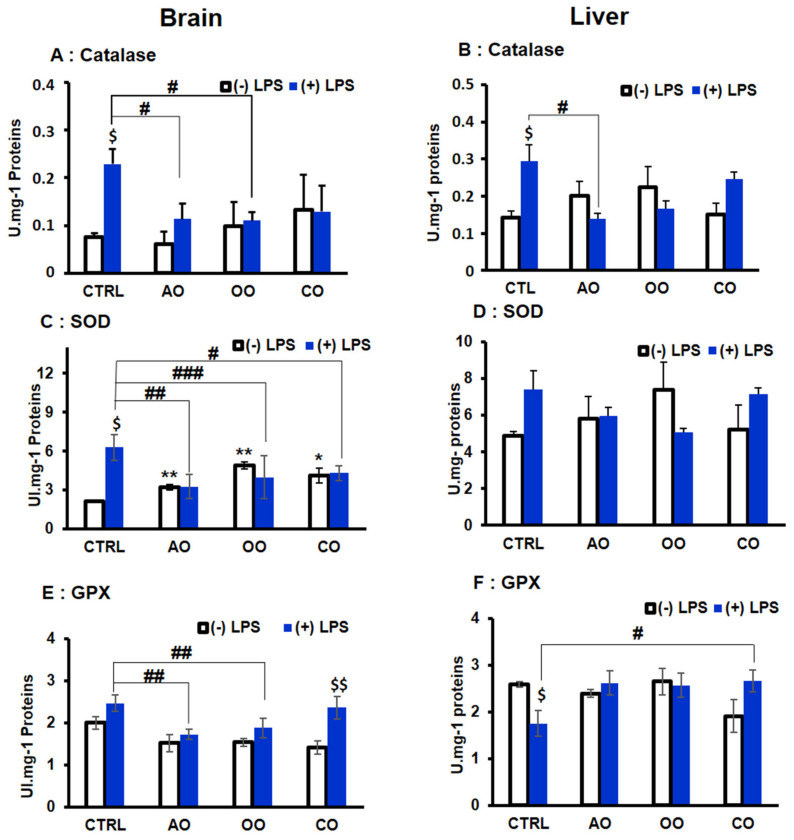
During sepsis, the imbalance between oxidative insult and body antioxidant response causes the dysfunction of organs, including the brain and liver. Exposing mice to bacterial lipopolysaccharides (LPS) results in a similar pathophysiological outcome. The protection offered by argan oil was studied against LPS-induced oxidative stress, dysregulation of peroxisomal antioxidants, and β-oxidation activities in the brain and liver. In a short-term LPS treatment, lipid peroxidation (malonaldehyde assay) increased in the brain and liver with upregulations of proinflammatory tumor necrosis factor (Tnf)-α and anti-inflammatory interleukin (Il)-10 genes, especially in the liver. Although exposure to olive oil (OO), colza oil (CO), and argan oil (AO) prevented LPS-induced lipid peroxidation in the brain and liver, only AO exposure protected against liver inflammation. Remarkably, only exposure to AO prevented LPS-dependent glutathione (GSH) dysregulation in the brain and liver. Furthermore, exposure to AO increased more efficiently than OO and CO in both organs, peroxisomal antioxidant capacity via induction of catalase (Cat) gene, protein and activity expression levels, and superoxide dismutase (Sod1) mRNA and activity levels. Interestingly, LPS decreased protein levels of the peroxisomal fatty acid-ATP binding cassette (ABC) transporters, ABCD1 and ABCD2, and increased acyl-CoA oxidase 1 (ACOX1) protein expression. Moreover, these LPS effects were attenuated for ABCD1 and ACOX1 in the brain of mice pretreated with AO. Our data collectively highlight the protective effects of AO against early oxidative stress caused by LPS in the brain and liver and their reliance on the preservation of peroxisomal functions, including antioxidant and β-oxidation activities, making AO a promising candidate for the prevention and management of sepsis.
Keywords: Acyl-CoA oxidase 1; LPS; antioxidant; argan oil; beta-oxidation; brain; catalase; glutathione peroxidase; peroxisome; superoxide dismutase.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
LinkOut - more resources
Full Text Sources
Miscellaneous
