

FK506-Binding Protein 11 Is a Novel Plasma Cell-Specific Antibody Folding Catalyst with Increased Expression in Idiopathic Pulmonary Fibrosis
- PMID: 35456020
- PMCID: PMC9027113
- DOI: 10.3390/cells11081341
FK506-Binding Protein 11 Is a Novel Plasma Cell-Specific Antibody Folding Catalyst with Increased Expression in Idiopathic Pulmonary Fibrosis
Abstract
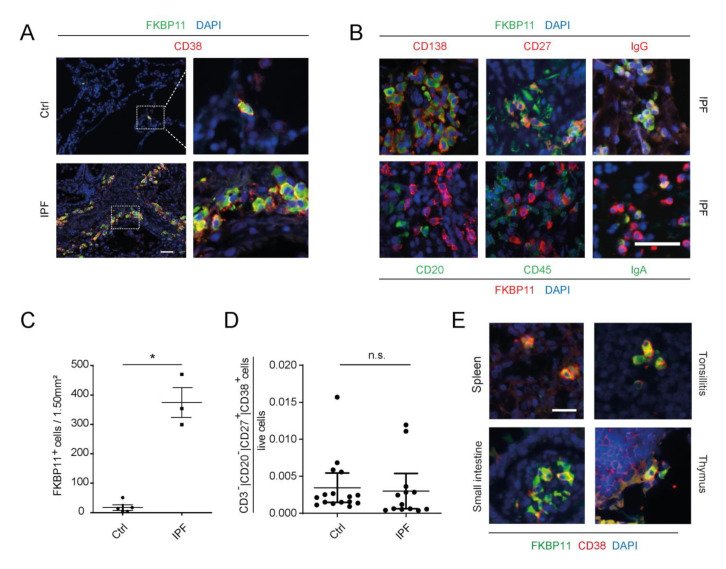
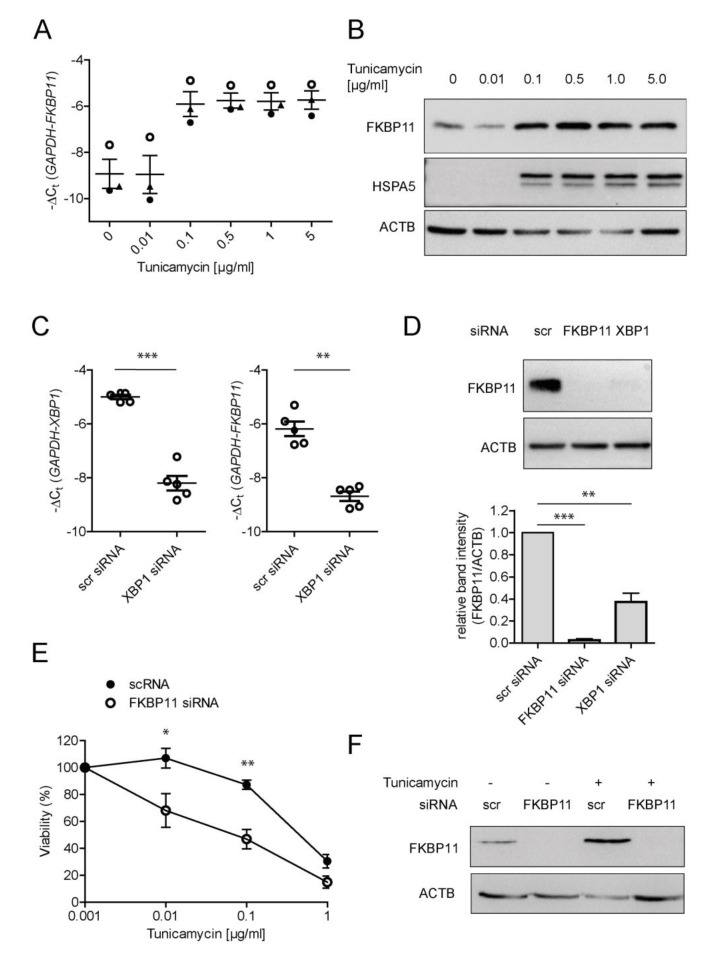
Antibodies are central effectors of the adaptive immune response, widespread used therapeutics, but also potentially disease-causing biomolecules. Antibody folding catalysts in the plasma cell are incompletely defined. Idiopathic pulmonary fibrosis (IPF) is a fatal chronic lung disease with increasingly recognized autoimmune features. We found elevated expression of FK506-binding protein 11 (FKBP11) in IPF lungs where FKBP11 specifically localized to antibody-producing plasma cells. Suggesting a general role in plasma cells, plasma cell-specific FKBP11 expression was equally observed in lymphatic tissues, and in vitro B cell to plasma cell differentiation was accompanied by induction of FKBP11 expression. Recombinant human FKBP11 was able to refold IgG antibody in vitro and inhibited by FK506, strongly supporting a function as antibody peptidyl-prolyl cis-trans isomerase. Induction of ER stress in cell lines demonstrated induction of FKBP11 in the context of the unfolded protein response in an X-box-binding protein 1 (XBP1)-dependent manner. While deficiency of FKBP11 increased susceptibility to ER stress-mediated cell death in an alveolar epithelial cell line, FKBP11 knockdown in an antibody-producing hybridoma cell line neither induced cell death nor decreased expression or secretion of IgG antibody. Similarly, antibody secretion by the same hybridoma cell line was not affected by knockdown of the established antibody peptidyl-prolyl isomerase cyclophilin B. The results are consistent with FKBP11 as a novel XBP1-regulated antibody peptidyl-prolyl cis-trans isomerase and indicate significant redundancy in the ER-resident folding machinery of antibody-producing hybridoma cells.
Keywords: ER stress; FK506-binding protein; antibody folding; immunophilin; interstitial lung disease; lung fibrosis; peptidyl-prolyl isomerase; tacrolimus.
Conflict of interest statement
N.K. (Naftali Kaminski) served as a consultant to Boehringer Ingelheim, Third Rock, Pliant, Samumed, NuMedii, Theravance, LifeMax, Three Lake Partners, Optikira, Astra Zeneca, RohBar, Veracyte, Augmanity, CSL Behring, Galapagos, Gilead and Thyron over the last 3 years, reports Equity in Pliant and Thyron, and a grant from Veracyte, Boehringer Ingelheim, BMS and non-financial support from MiRagen and Astra Zeneca. N.K. (Naftali Kaminski) has IP on novel biomarkers and therapeutics in IPF licensed to Biotech. E.M. received honorarium from Roche, Novartis, Merck, Sanofi, Biogen, and Bioeq and grant support from Novartis, Sanofi, Roche and Merck. O.E. is supported by Bristol Myers Squibb (FP00018992) and serves in advisory capacity to Pieris Pharmaceuticals, Blade Therapeutics, Delta 4 and YAP Therapeutics. L.K. is employee of Secarna Pharmaceuticals, but was still Ph.D. candidate in Staab-Weijnitz Lab when she contributed to the study. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results. All other authors declare no conflict of interest.
Figures




References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
