

Conformational transitions in BTG1 antiproliferative protein and their modulation by disease mutants
- PMID: 35459639
- PMCID: PMC9617077
- DOI: 10.1016/j.bpj.2022.04.023
Conformational transitions in BTG1 antiproliferative protein and their modulation by disease mutants
Abstract
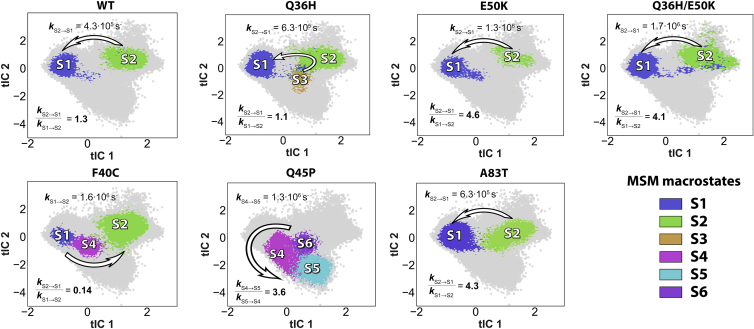
B cell translocation gene 1 (BTG1) protein belongs to the BTG/transducer of ERBB2 (TOB) family of antiproliferative proteins whose members regulate various key cellular processes such as cell cycle progression, apoptosis, and differentiation. Somatic missense mutations in BTG1 are found in ∼70% of a particularly malignant and disseminated subtype of diffuse large B cell lymphoma (DLBCL). Antiproliferative activity of BTG1 has been linked to its ability to associate with transcriptional cofactors and various enzymes. However, molecular mechanisms underlying these functional interactions and how the disease-linked mutations in BTG1 affect these mechanisms are currently unknown. To start filling these knowledge gaps, here, using atomistic molecular dynamics (MD) simulations, we explored structural, dynamic, and kinetic characteristics of BTG1 protein, and studied how various DLBCL mutations affect these characteristics. We focused on the protein region formed by α2 and α4 helices, as this interface has been reported not only to serve as a binding hotspot for several cellular partners but also to harbor sites for the majority of known DLBCL mutations. Markov state modeling analysis of extensive MD simulations revealed that the α2-α4 interface in the wild-type (WT) BTG1 undergoes conformational transitions between closed and open metastable states. Importantly, we show that some of the mutations in this region that are observed in DLBCL, such as Q36H, F40C, Q45P, E50K (in α2), and A83T and A84E (in α4), either overstabilize one of these two metastable states or give rise to new conformations in which these helices are distorted (i.e., kinked or unfolded). Based on these results, we conclude that the rapid interconversion between the closed and open conformations of the α2-α4 interface is an essential component of the BTG1 functional dynamics that can prime the protein for functional associations with its binding partners. Disruption of the native dynamic equilibrium by DLBCL mutants leads to the ensemble of conformations in BTG1 that are unlikely structurally and/or kinetically to enable productive functional interactions with the binding proteins.
Copyright © 2022 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests A.M. has research funding from Janssen Pharmaceuticals, Sanofi, Epizyme, and Daiichi Sankyo; has consulted for Epizyme and Constellation; and is on the advisory board for KDAC Pharma.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
