

Extracellular Nucleotides and Histamine Suppress TLR3- and RIG-I-Mediated Release of Antiviral IFNs from Human Airway Epithelial Cells
- PMID: 35459743
- PMCID: PMC9444327
- DOI: 10.4049/jimmunol.2101085
Extracellular Nucleotides and Histamine Suppress TLR3- and RIG-I-Mediated Release of Antiviral IFNs from Human Airway Epithelial Cells
Abstract
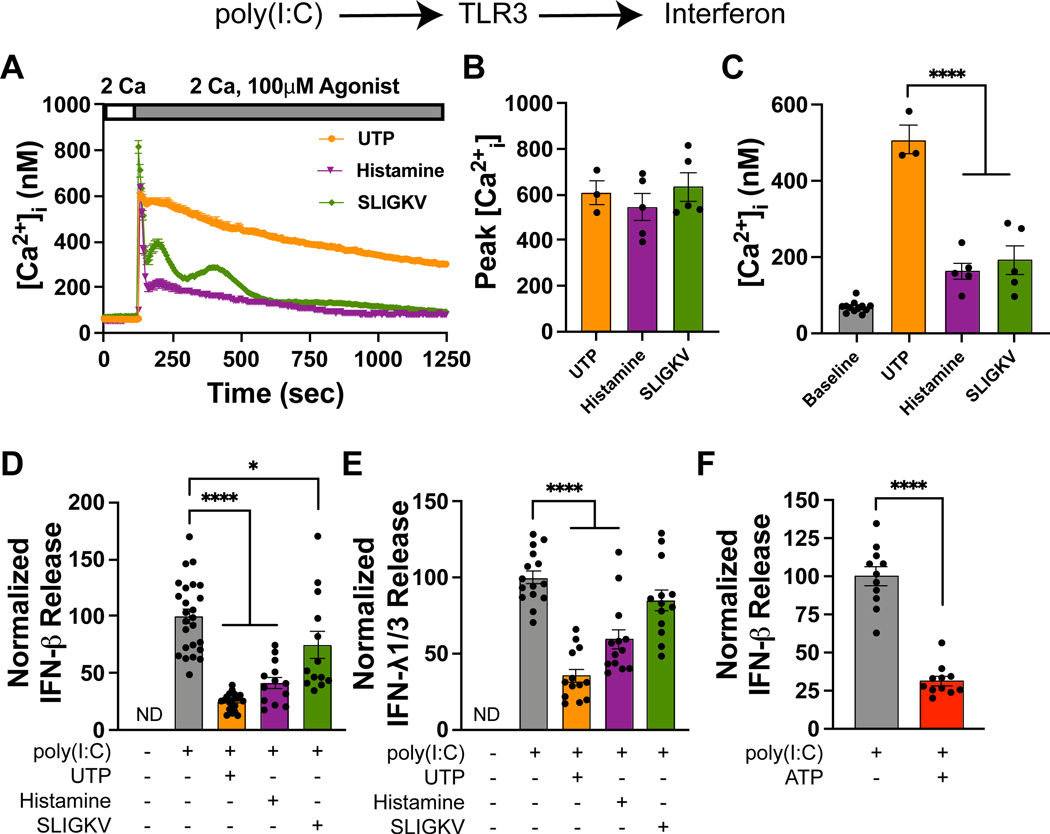
Respiratory viruses stimulate the release of antiviral IFNs from the airway epithelium. Previous studies have shown that asthmatic patients show diminished release of type I and type III IFNs from bronchial epithelia. However, the mechanism of this suppression is not understood. In this study, we report that extracellular nucleotides and histamine, which are elevated in asthmatic airways, strongly inhibit release of type I and type III IFNs from human bronchial airway epithelial cells (AECs). Specifically, ATP, UTP, and histamine all inhibited the release of type I and type III IFNs from AECs induced by activation of TLR3, retinoic acid-inducible gene I (RIG-I), or cyclic GMP-AMP synthase-STING. This inhibition was at least partly mediated by Gq signaling through purinergic P2Y2 and H1 receptors, but it did not involve store-operated calcium entry. Pharmacological blockade of protein kinase C partially reversed inhibition of IFN production. Conversely, direct activation of protein kinase C with phorbol esters strongly inhibited TLR3- and RIG-I-mediated IFN production. Inhibition of type I and type III IFNs by ATP, UTP, histamine, and the proteinase-activated receptor 2 (PAR2) receptor agonist SLIGKV also occurred in differentiated AECs grown at an air-liquid interface, indicating that the suppression is conserved following mucociliary differentiation. Importantly, histamine and, more strikingly, ATP inhibited type I IFN release from human airway cells infected with live influenza A virus or rhinovirus 1B. These results reveal an important role for extracellular nucleotides and histamine in attenuating the induction of type I and III IFNs from AECs and help explain the molecular basis of the suppression of IFN responses in asthmatic patients.
Copyright © 2022 by The American Association of Immunologists, Inc.
Conflict of interest statement
Competing interests
N/A.
Figures





References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
