

Structure and mechanism of the γ-secretase intramembrane protease complex
- PMID: 35461161
- PMCID: PMC9189058
- DOI: 10.1016/j.sbi.2022.102373
Structure and mechanism of the γ-secretase intramembrane protease complex
Abstract
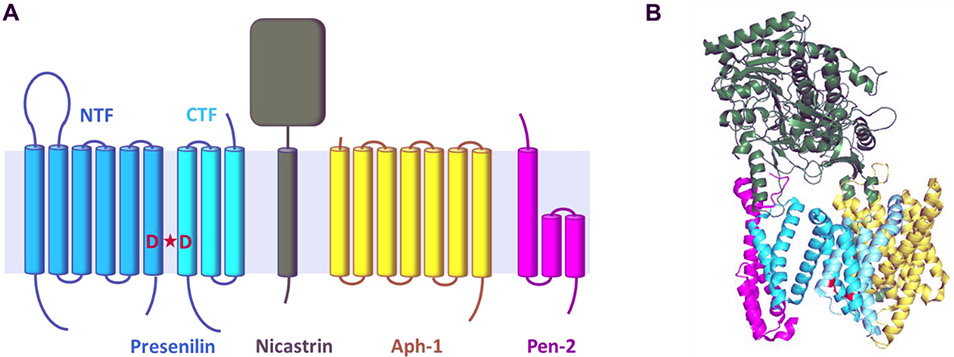
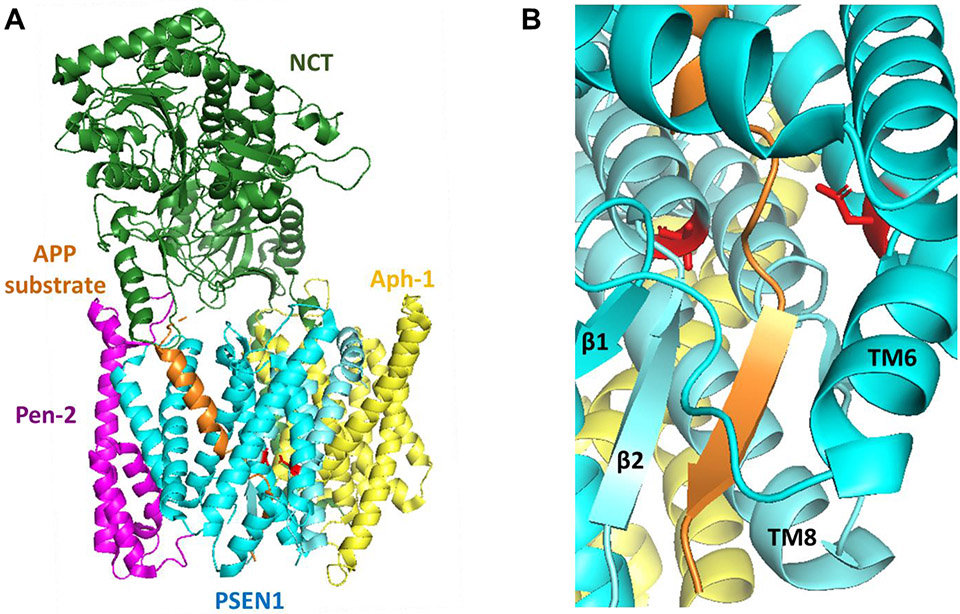
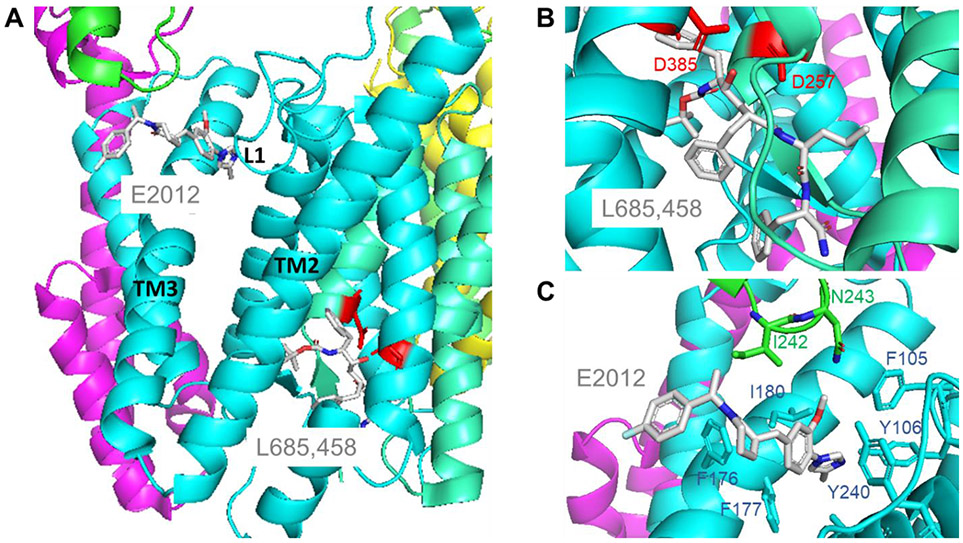
γ-Secretase is a membrane protein complex that proteolyzes within the transmembrane domain of >100 substrates, including those derived from the amyloid precursor protein and the Notch family of cell surface receptors. The nine-transmembrane presenilin is the catalytic component of this aspartyl protease complex that carries out hydrolysis in the lipid bilayer. Advances in cryoelectron microscopy have led to the elucidation of the structure of the γ-secretase complex at atomic resolution. Recently, structures of the enzyme have been determined with bound APP- or Notch-derived substrates, providing insight into the nature of substrate recognition and processing. Molecular dynamics simulations of substrate-bound enzymes suggest dynamic mechanisms of intramembrane proteolysis. Structures of the enzyme bound to small-molecule inhibitors and modulators have also been solved, setting the stage for rational structure-based drug discovery targeting γ-secretase.
Copyright © 2022 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Conflicts statement
Nothing to declare.
Figures

Similar articles
-
Molecular Dynamics Activation of γ-Secretase for Cleavage of the Notch1 Substrate.ACS Chem Neurosci. 2023 Dec 6;14(23):4216-4226. doi: 10.1021/acschemneuro.3c00594. Epub 2023 Nov 9. ACS Chem Neurosci. 2023. PMID: 37942767 Free PMC article.
-
Recognition of the amyloid precursor protein by human γ-secretase.Science. 2019 Feb 15;363(6428):eaaw0930. doi: 10.1126/science.aaw0930. Epub 2019 Jan 10. Science. 2019. PMID: 30630874
-
Structure and Function of the γ-Secretase Complex.Biochemistry. 2019 Jul 9;58(27):2953-2966. doi: 10.1021/acs.biochem.9b00401. Epub 2019 Jun 25. Biochemistry. 2019. PMID: 31198028 Free PMC article.
-
Substrate recognition and processing by γ-secretase.Biochim Biophys Acta Biomembr. 2020 Jan 1;1862(1):183016. doi: 10.1016/j.bbamem.2019.07.004. Epub 2019 Jul 8. Biochim Biophys Acta Biomembr. 2020. PMID: 31295475 Free PMC article. Review.
-
Structure, mechanism and inhibition of gamma-secretase and presenilin-like proteases.Biol Chem. 2010 Aug;391(8):839-47. doi: 10.1515/BC.2010.086. Biol Chem. 2010. PMID: 20482315 Free PMC article. Review.
Cited by
-
Comprehensive Catalog of Variants Potentially Associated with Hidradenitis Suppurativa, Including Newly Identified Variants from a Cohort of 100 Patients.Int J Mol Sci. 2024 Sep 26;25(19):10374. doi: 10.3390/ijms251910374. Int J Mol Sci. 2024. PMID: 39408704 Free PMC article.
-
Maintaining the Integral Membrane Proteome: Revisiting the Functional Repertoire of Integral Membrane Proteases.Chembiochem. 2025 May 5;26(9):e202500048. doi: 10.1002/cbic.202500048. Epub 2025 Mar 18. Chembiochem. 2025. PMID: 40056010 Free PMC article. Review.
-
Interactions between amyloid, amyloid precursor protein, and mitochondria.Biochem Soc Trans. 2023 Feb 27;51(1):173-182. doi: 10.1042/BST20220518. Biochem Soc Trans. 2023. PMID: 36688439 Free PMC article.
-
Effects of presenilin-1 familial Alzheimer's disease mutations on γ-secretase activation for cleavage of amyloid precursor protein.Commun Biol. 2023 Feb 14;6(1):174. doi: 10.1038/s42003-023-04539-1. Commun Biol. 2023. PMID: 36788318 Free PMC article.
-
Clinical significance of PNO1 as a novel biomarker and therapeutic target of hepatocellular carcinoma.J Cell Mol Med. 2024 May;28(9):e18295. doi: 10.1111/jcmm.18295. J Cell Mol Med. 2024. PMID: 38722284 Free PMC article.
References
-
- Sun L, Li X, Shi Y: Structural biology of intramembrane proteases: mechanistic insights from rhomboid and S2P to γ-secretase. Curr Opin Struct Biol 2016, 37:97–107. - PubMed
-
- Wolfe MS, Xia W, Ostaszewski BL, Diehl TS, Kimberly WT, Selkoe DJ: Two transmembrane aspartates in presenilin-1 required for presenilin endoproteolysis and γ-secretase activity. Nature 1999, 398:513–517. - PubMed
-
- De Strooper B: Aph-1, Pen-2, and nicastrin with presenilin generate an active γ-secretase complex. Neuron 2003, 38:9–12. - PubMed
-
- Thinakaran G, Borchelt DR, Lee MK, Slunt HH, Spitzer L, Kim G, Ratovitsky T, Davenport F, Nordstedt C, Seeger M, et al.: Endoproteolysis of presenilin 1 and accumulation of processed derivatives in vivo. Neuron 1996, 17:181–190. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
