

ZmPP2C26 Alternative Splicing Variants Negatively Regulate Drought Tolerance in Maize
- PMID: 35463404
- PMCID: PMC9024303
- DOI: 10.3389/fpls.2022.851531
ZmPP2C26 Alternative Splicing Variants Negatively Regulate Drought Tolerance in Maize
Abstract
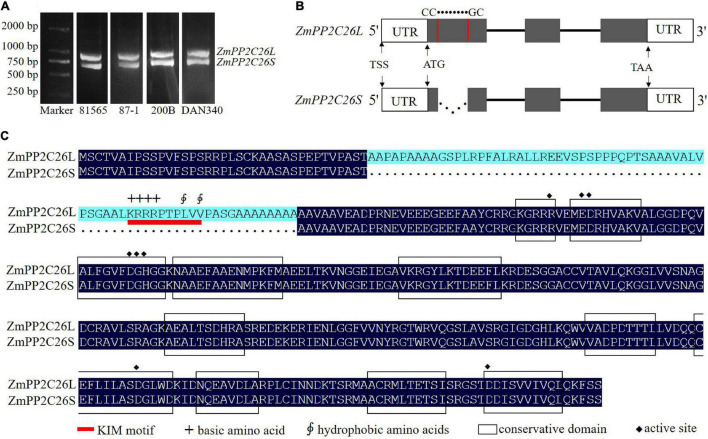
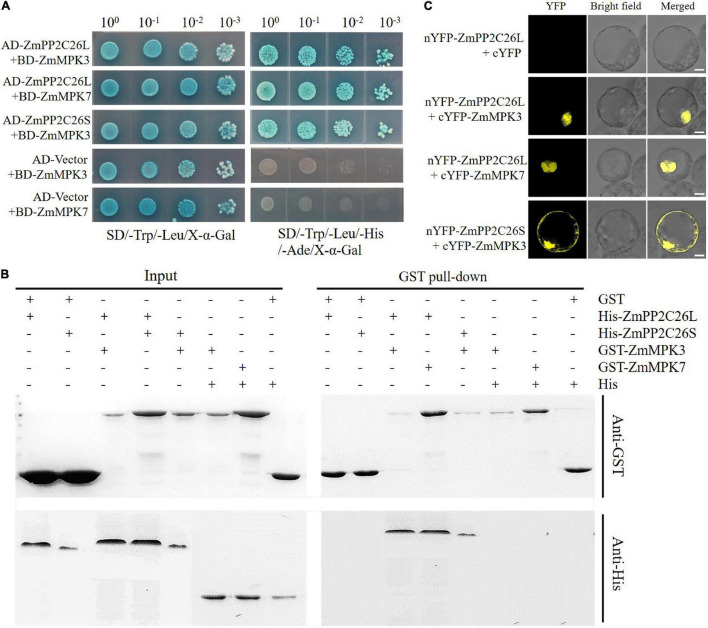
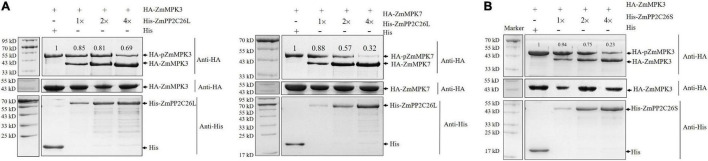
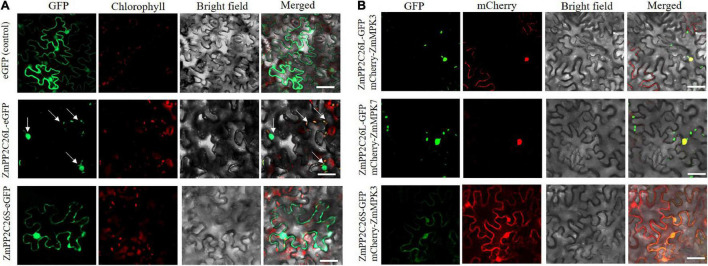
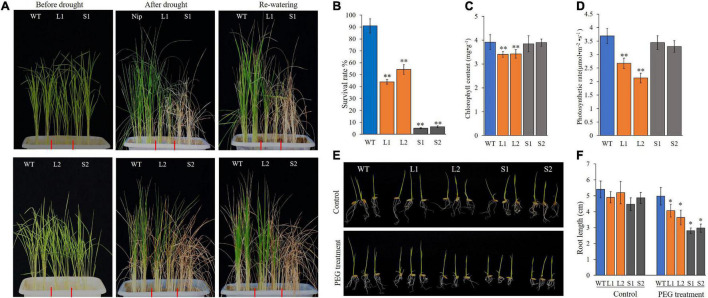
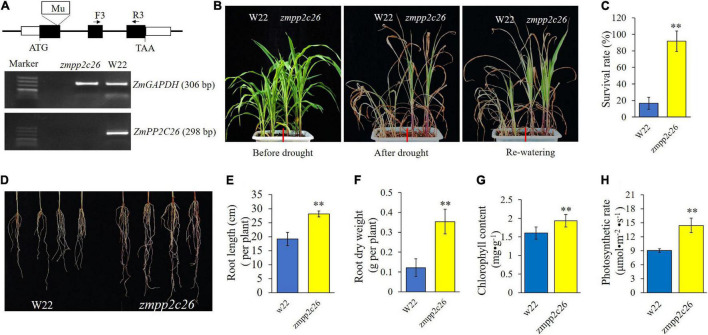
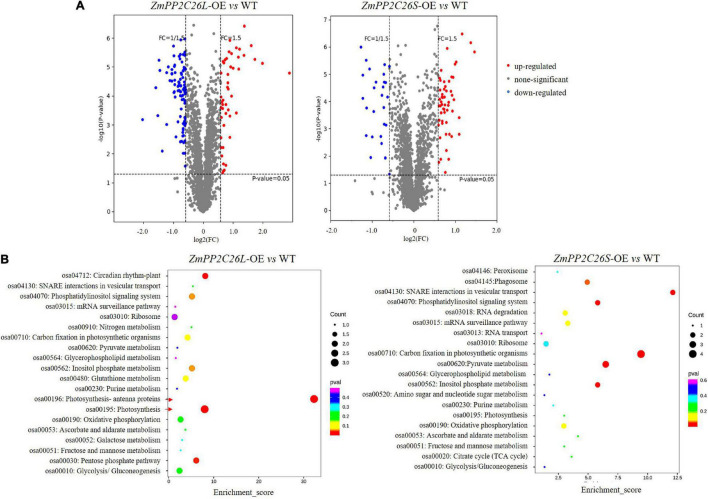
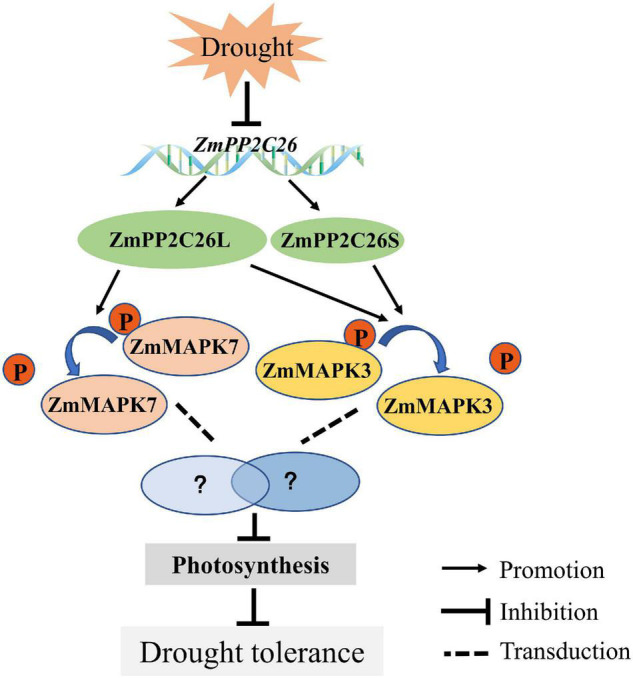
Serine/threonine protein phosphatase 2C (PP2C) dephosphorylates proteins and plays crucial roles in plant growth, development, and stress response. In this study, we characterized a clade B member of maize PP2C family, i.e., ZmPP2C26, that negatively regulated drought tolerance by dephosphorylating ZmMAPK3 and ZmMAPK7 in maize. The ZmPP2C26 gene generated ZmPP2C26L and ZmPP2C26S isoforms through untypical alternative splicing. ZmPP2C26S lost 71 amino acids including an MAPK interaction motif and showed higher phosphatase activity than ZmPP2C26L. ZmPP2C26L directly interacted with, dephosphorylated ZmMAPK3 and ZmMAPK7, and localized in chloroplast and nucleus, but ZmPP2C26S only dephosphorylated ZmMAPK3 and localized in cytosol and nucleus. The expression of ZmPP2C26L and ZmPP2C26 was significantly inhibited by drought stress. Meanwhile, the maize zmpp2c26 mutant exhibited enhancement of drought tolerance with higher root length, root weight, chlorophyll content, and photosynthetic rate compared with wild type. However, overexpression of ZmPP2C26L and ZmPP2C26S significantly decreased drought tolerance in Arabidopsis and rice with lower root length, chlorophyll content, and photosynthetic rate. Phosphoproteomic analysis revealed that the ZmPP2C26 protein also altered phosphorylation level of proteins involved in photosynthesis. This study provides insights into understanding the mechanism of PP2C in response to abiotic stress.
Keywords: MAPK; alternative splicing; drought stress; maize; photosynthesis; protein phosphatase 2C.
Copyright © 2022 Lu, Li, Peng, Cao, Qu, Sun, Yang, Lu, Zhang, Zheng, Fu and Yu.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
Isolation and characterization of maize ZmPP2C26 gene promoter in drought-response.Physiol Mol Biol Plants. 2020 Nov;26(11):2189-2197. doi: 10.1007/s12298-020-00910-2. Epub 2020 Nov 18. Physiol Mol Biol Plants. 2020. PMID: 33268922 Free PMC article.
-
Identification of the Maize PP2C Gene Family and Functional Studies on the Role of ZmPP2C15 in Drought Tolerance.Plants (Basel). 2024 Jan 23;13(3):340. doi: 10.3390/plants13030340. Plants (Basel). 2024. PMID: 38337873 Free PMC article.
-
Deletion of an Endoplasmic Reticulum Stress Response Element in a ZmPP2C-A Gene Facilitates Drought Tolerance of Maize Seedlings.Mol Plant. 2017 Mar 6;10(3):456-469. doi: 10.1016/j.molp.2016.10.003. Epub 2016 Oct 13. Mol Plant. 2017. PMID: 27746300
-
ABA inducible rice protein phosphatase 2C confers ABA insensitivity and abiotic stress tolerance in Arabidopsis.PLoS One. 2015 Apr 17;10(4):e0125168. doi: 10.1371/journal.pone.0125168. eCollection 2015. PLoS One. 2015. PMID: 25886365 Free PMC article.
-
Transcriptional regulatory networks in response to drought stress and rewatering in maize (Zea mays L.).Mol Genet Genomics. 2021 Nov;296(6):1203-1219. doi: 10.1007/s00438-021-01820-y. Epub 2021 Oct 3. Mol Genet Genomics. 2021. PMID: 34601650 Review.
Cited by
-
Transcription Factor ZmNAC20 Improves Drought Resistance by Promoting Stomatal Closure and Activating Expression of Stress-Responsive Genes in Maize.Int J Mol Sci. 2023 Mar 1;24(5):4712. doi: 10.3390/ijms24054712. Int J Mol Sci. 2023. PMID: 36902144 Free PMC article.
-
Marker-Assisted Backcross Breeding of Drought-Tolerant Maize Lines Transformed by Vacuolar H+-Pyrophosphatase Gene (AnVP1) from Ammopiptanthus nanus.Plants (Basel). 2025 Mar 15;14(6):926. doi: 10.3390/plants14060926. Plants (Basel). 2025. PMID: 40265817 Free PMC article.
-
Genome-Wide Identification and Expression Analysis of PP2C Gene Family in Eelgrass.Genes (Basel). 2025 May 29;16(6):657. doi: 10.3390/genes16060657. Genes (Basel). 2025. PMID: 40565549 Free PMC article.
-
Genome-Wide Identification, Expression and Interaction Analyses of PP2C Family Genes in Chenopodium quinoa.Genes (Basel). 2023 Dec 27;15(1):41. doi: 10.3390/genes15010041. Genes (Basel). 2023. PMID: 38254931 Free PMC article.
-
Interaction between SlMAPK3 and SlASR4 regulates drought resistance in tomato (Solanum lycopersicum L.).Mol Breed. 2023 Oct 3;43(10):73. doi: 10.1007/s11032-023-01418-9. eCollection 2023 Oct. Mol Breed. 2023. PMID: 37795156 Free PMC article.
References
-
- Brock A. K., Willmann R., Kolb D., Grefen L., Lajunen H. M., Bethke G., et al. (2010). The Arabidopsis mitogen-activated protein kinase phosphatase PP2C5 affects seed germination, stomatal aperture, and abscisic acid-inducible gene expression. Plant Physiol. 153 1098–1111. 10.1104/pp.110.156109 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources