

In vivo Functional Characterization of Hydrophilic X2 Modules in the Cellulosomal Scaffolding Protein
- PMID: 35464986
- PMCID: PMC9022034
- DOI: 10.3389/fmicb.2022.861549
In vivo Functional Characterization of Hydrophilic X2 Modules in the Cellulosomal Scaffolding Protein
Abstract
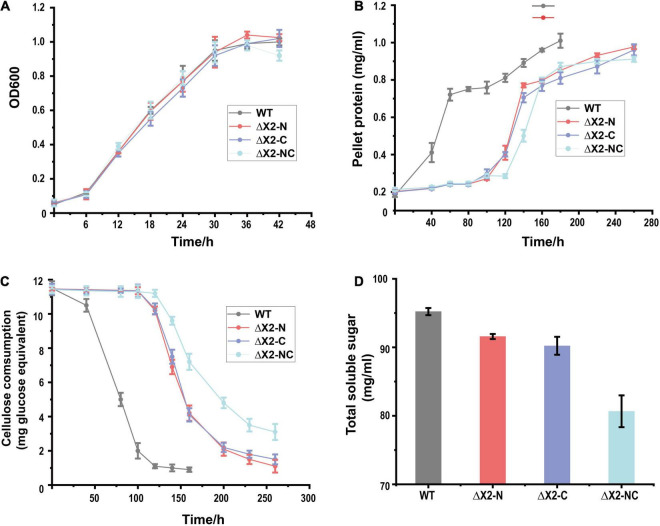
As part of free cellulases or scaffolding proteins in cellulosomes, the hydrophilic non-catalytic X2 module is widely distributed in cellulolytic Clostridia or other Firmicutes bacteria. Previous biochemical studies suggest that X2 modules might increase the solubility and substrate binding affinity of X2-bearing proteins. However, their in vivo biological functions remain elusive. Here we employed CRISPR-Cas9 editing to genetically modify X2 modules by deleting the conserved motif (NGNT) from the CipC scaffoldin. Both single and double X2 mutants (X2-N: near the N terminus of CipC; X2-C: near the C terminus of CipC) presented similar stoichiometric compositions in isolated cellulosomes as the wildtype strain (WT). These X2 mutants had an elongated adaptation stage during growth on cellulose compared to cellobiose. Compared to WT, the double mutant ΔX2-NC reduced cellulose degradation by 15% and the amount of released soluble sugars by 63%. Since single X2 mutants did not present such obvious physiological changes as ΔX2-NC, there seems to be a functional redundancy between X2 modules in CipC. The in vivo adhesion assay revealed that ΔX2-NC decreased cell attachment to cellulose by 70% but a weaker effect was also overserved in single X2 mutants. These results highlight the in vivo biological role of X2 in increasing cellulose degradation efficiency by enhancing the binding affinity between cells and cellulose, which provides new perspectives for microbial engineering.
Keywords: Clostridium cellulolyticum; X2 module; cellulose degradation; cellulosome; motif deletion.
Copyright © 2022 Tao, Liu, Kempher, Xu and Zhou.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



Similar articles
-
Are cellulosome scaffolding protein CipC and CBM3-containing protein HycP, involved in adherence of Clostridium cellulolyticum to cellulose?PLoS One. 2013 Jul 25;8(7):e69360. doi: 10.1371/journal.pone.0069360. Print 2013. PLoS One. 2013. PMID: 23935995 Free PMC article.
-
Turning a potent family-9 free cellulase into an operational cellulosomal component and vice versa.FEBS J. 2019 Sep;286(17):3359-3373. doi: 10.1111/febs.14858. Epub 2019 May 3. FEBS J. 2019. PMID: 31004451
-
Stoichiometric Assembly of the Cellulosome Generates Maximum Synergy for the Degradation of Crystalline Cellulose, as Revealed by In Vitro Reconstitution of the Clostridium thermocellum Cellulosome.Appl Environ Microbiol. 2015 Jul;81(14):4756-66. doi: 10.1128/AEM.00772-15. Epub 2015 May 8. Appl Environ Microbiol. 2015. PMID: 25956772 Free PMC article.
-
Cellulosomes-structure and ultrastructure.J Struct Biol. 1998 Dec 15;124(2-3):221-34. doi: 10.1006/jsbi.1998.4065. J Struct Biol. 1998. PMID: 10049808 Review.
-
Cellulosomes: Highly Efficient Cellulolytic Complexes.Subcell Biochem. 2021;96:323-354. doi: 10.1007/978-3-030-58971-4_9. Subcell Biochem. 2021. PMID: 33252735 Review.
Cited by
-
A genomic analysis reveals the diversity of cellulosome displaying bacteria.Front Microbiol. 2024 Oct 30;15:1473396. doi: 10.3389/fmicb.2024.1473396. eCollection 2024. Front Microbiol. 2024. PMID: 39539715 Free PMC article.
References
LinkOut - more resources
Full Text Sources
Research Materials