

Role of the gut-brain axis in energy and glucose metabolism
- PMID: 35474341
- PMCID: PMC9076644
- DOI: 10.1038/s12276-021-00677-w
Role of the gut-brain axis in energy and glucose metabolism
Abstract
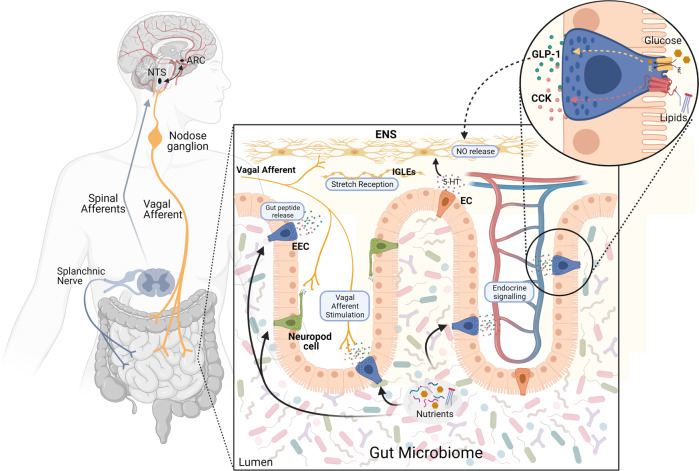
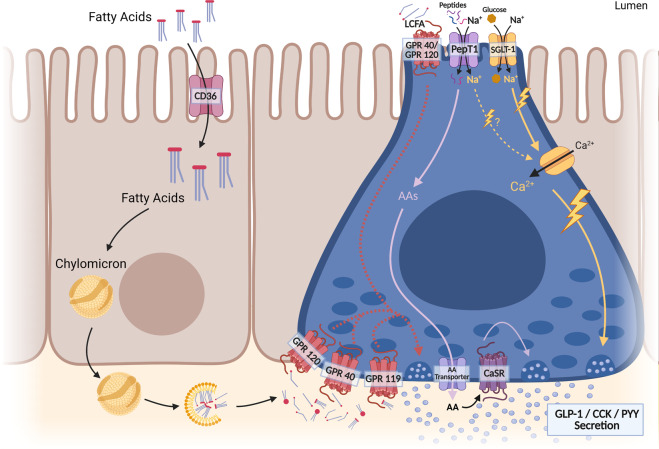
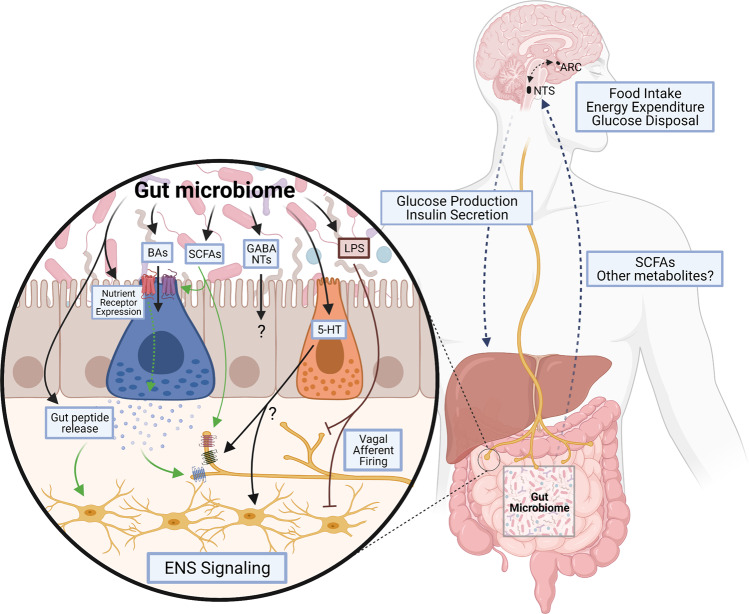
The gastrointestinal tract plays a role in the development and treatment of metabolic diseases. During a meal, the gut provides crucial information to the brain regarding incoming nutrients to allow proper maintenance of energy and glucose homeostasis. This gut-brain communication is regulated by various peptides or hormones that are secreted from the gut in response to nutrients; these signaling molecules can enter the circulation and act directly on the brain, or they can act indirectly via paracrine action on local vagal and spinal afferent neurons that innervate the gut. In addition, the enteric nervous system can act as a relay from the gut to the brain. The current review will outline the different gut-brain signaling mechanisms that contribute to metabolic homeostasis, highlighting the recent advances in understanding these complex hormonal and neural pathways. Furthermore, the impact of the gut microbiota on various components of the gut-brain axis that regulates energy and glucose homeostasis will be discussed. A better understanding of the gut-brain axis and its complex relationship with the gut microbiome is crucial for the development of successful pharmacological therapies to combat obesity and diabetes.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Hales, C. M., Carroll, M. D., Fryar, C. D. & Ogden, C. L. Prevalence of obesity and severe obesity among adults: United States, 2017–2018. NCHS Data Brief, 1–8 (2020). - PubMed
-
- Reid T. Choosing GLP-1 receptor agonists or DPP-4 inhibitors: weighing the clinical trial evidence. Clin. Diabetes. 2012;30:3. doi: 10.2337/diaclin.30.1.3. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
