

TAU ablation in excitatory neurons and postnatal TAU knockdown reduce epilepsy, SUDEP, and autism behaviors in a Dravet syndrome model
- PMID: 35476595
- PMCID: PMC9102397
- DOI: 10.1126/scitranslmed.abm5527
TAU ablation in excitatory neurons and postnatal TAU knockdown reduce epilepsy, SUDEP, and autism behaviors in a Dravet syndrome model
Abstract
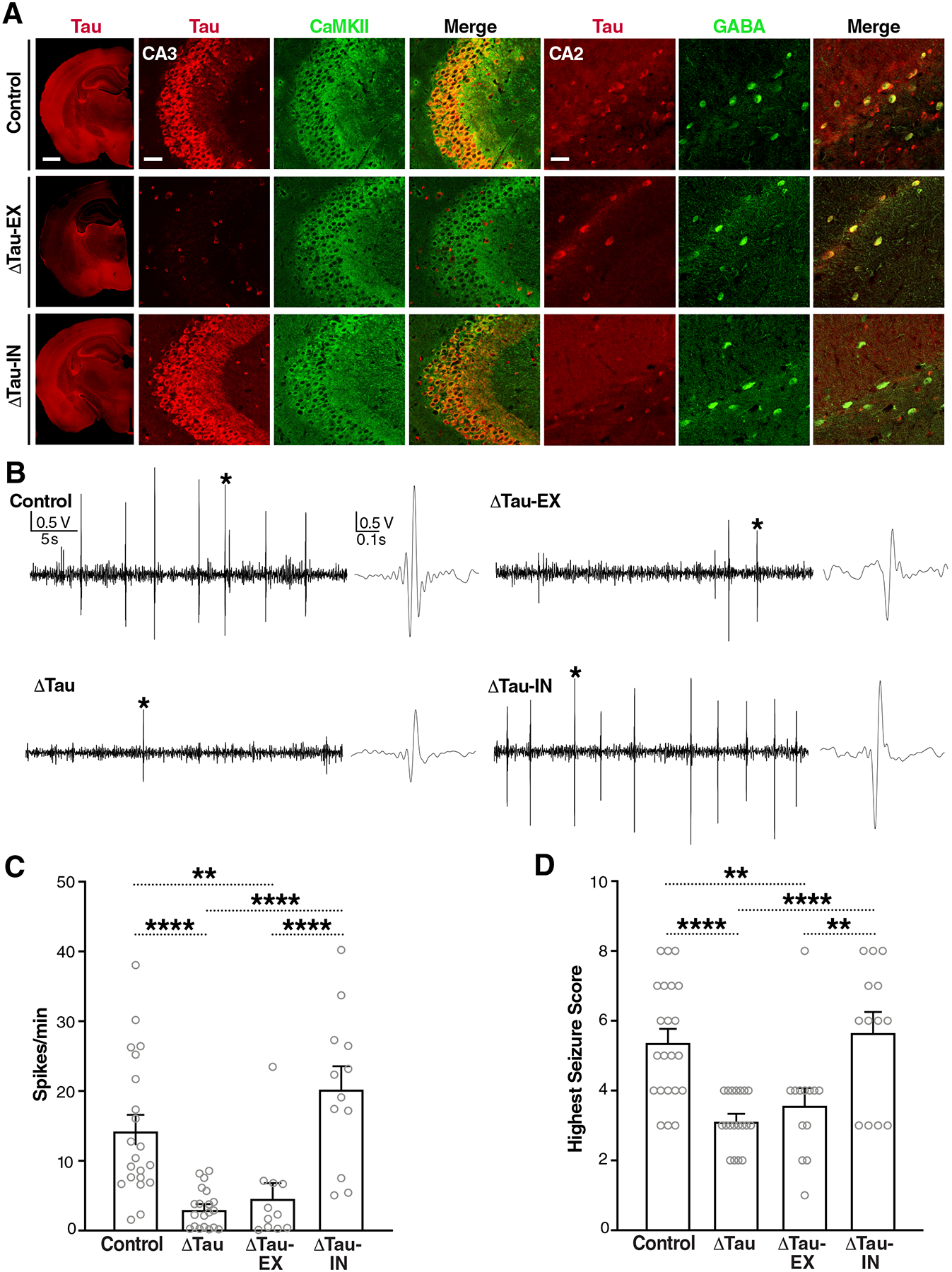
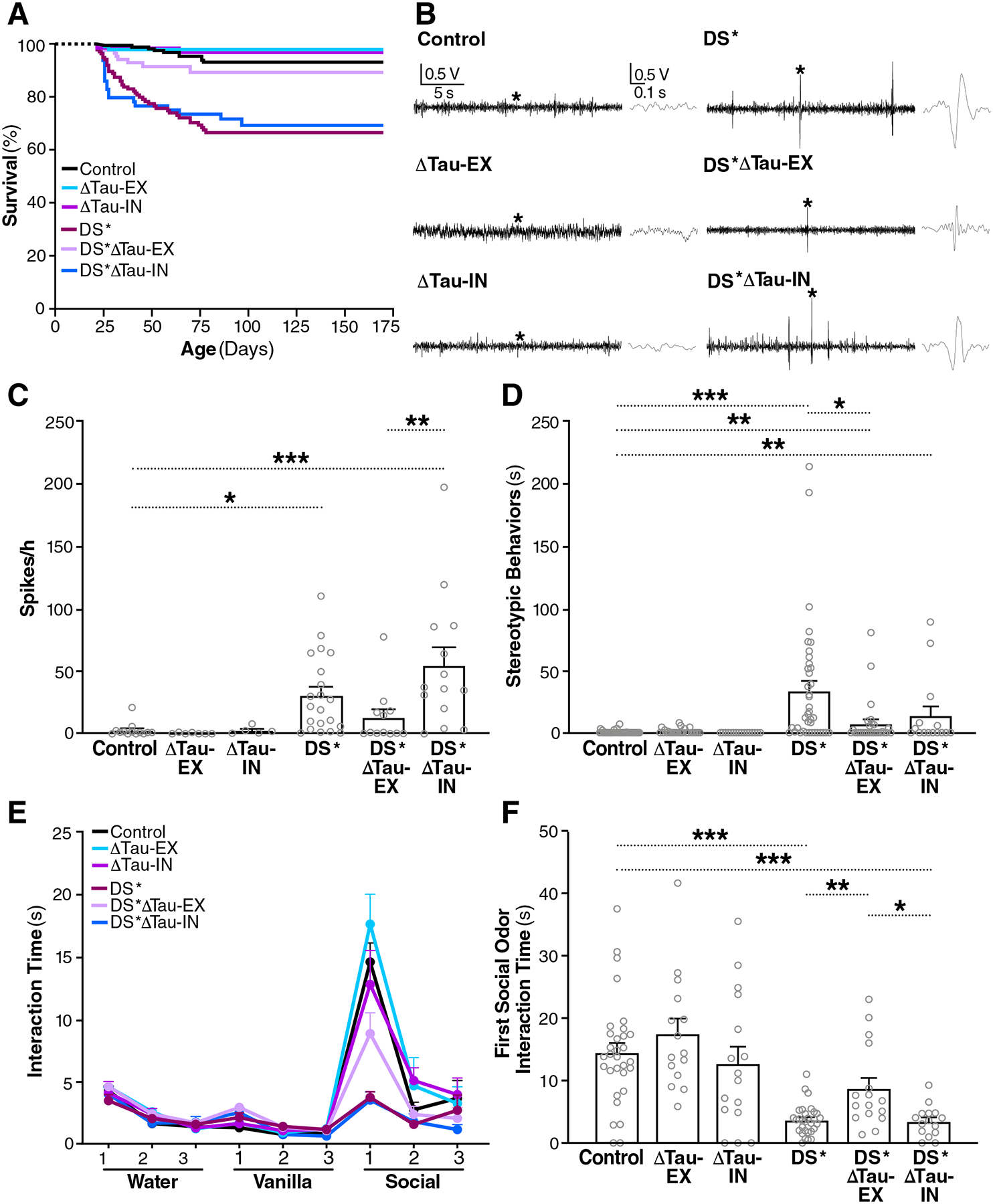
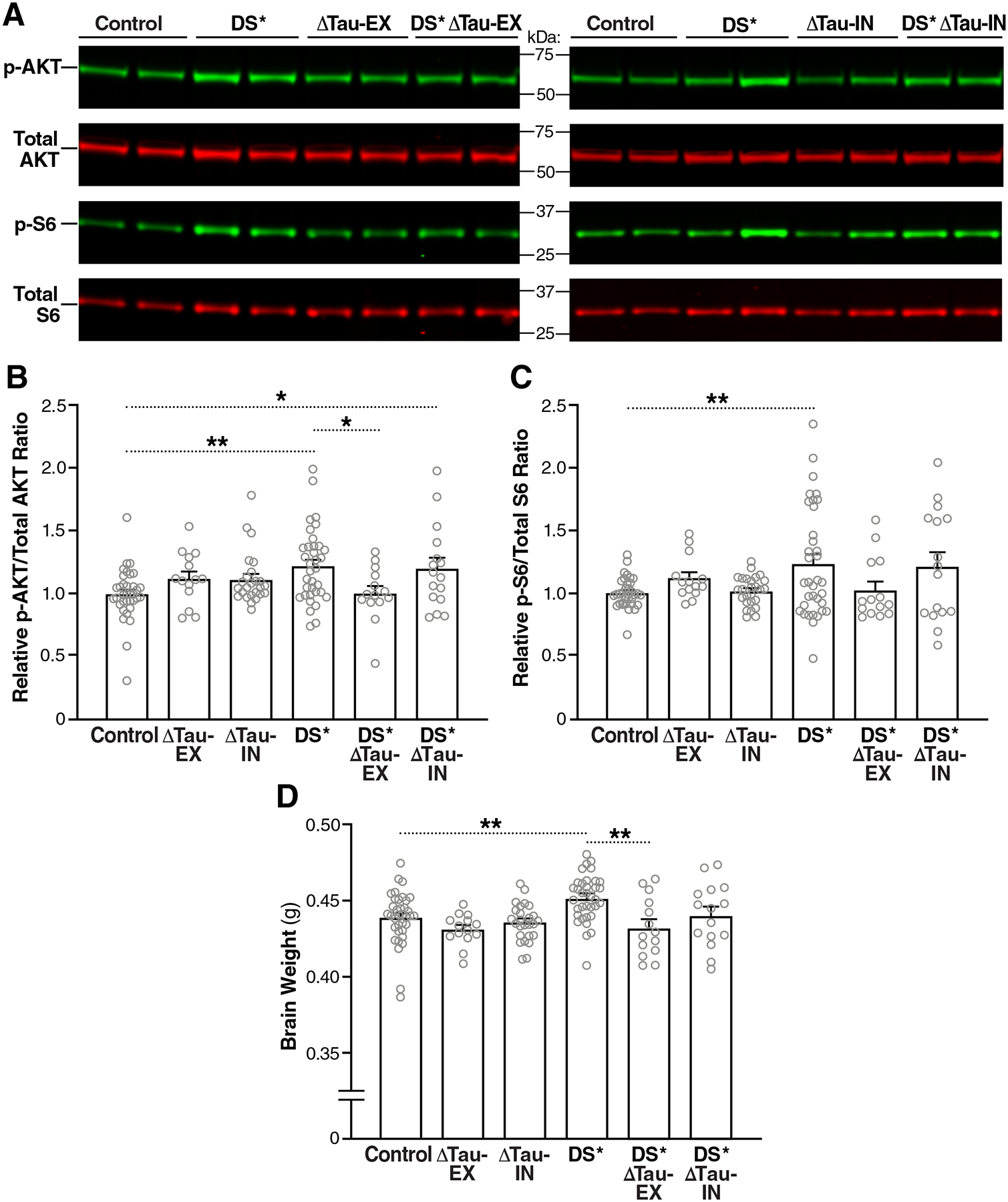
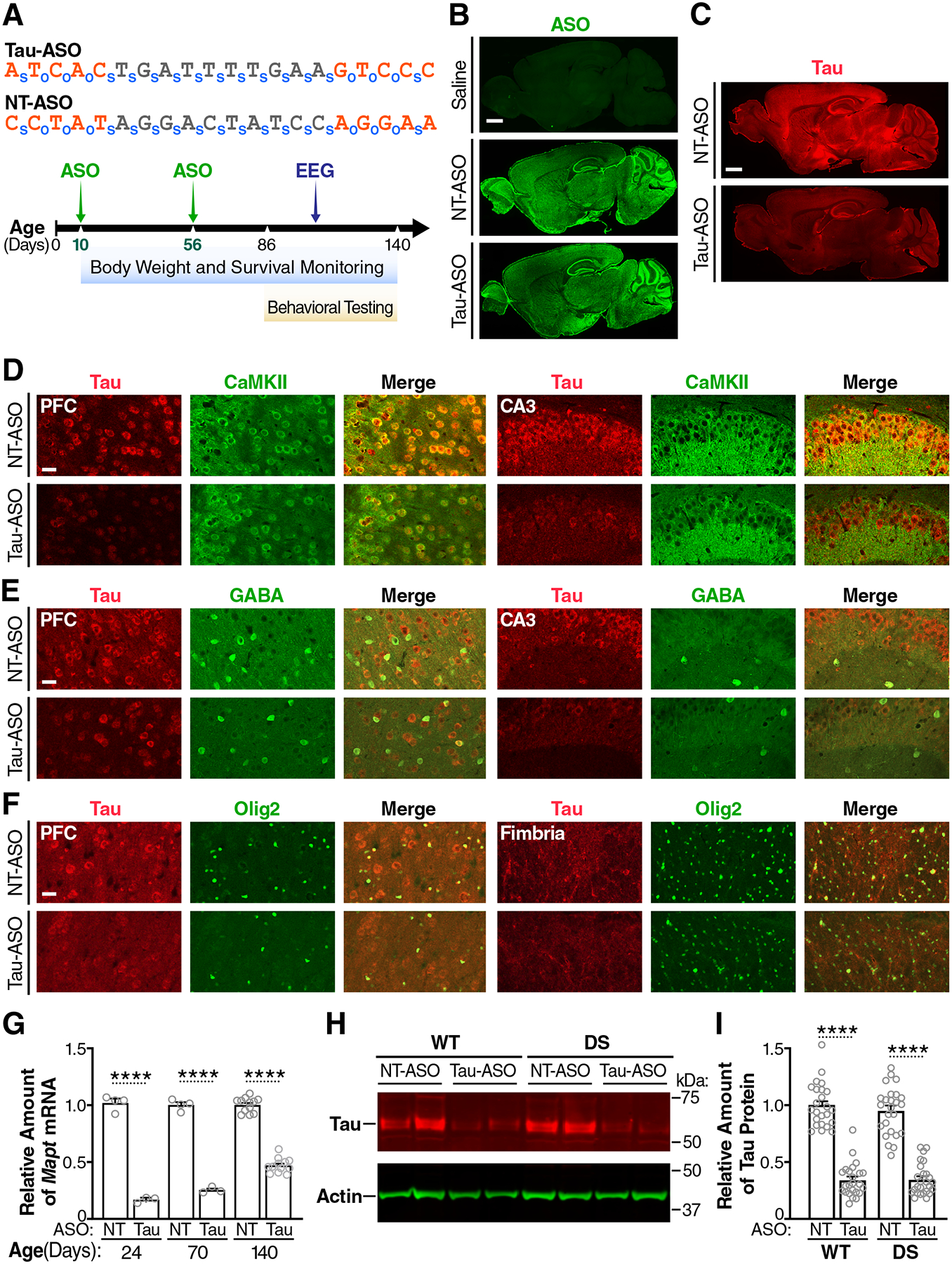
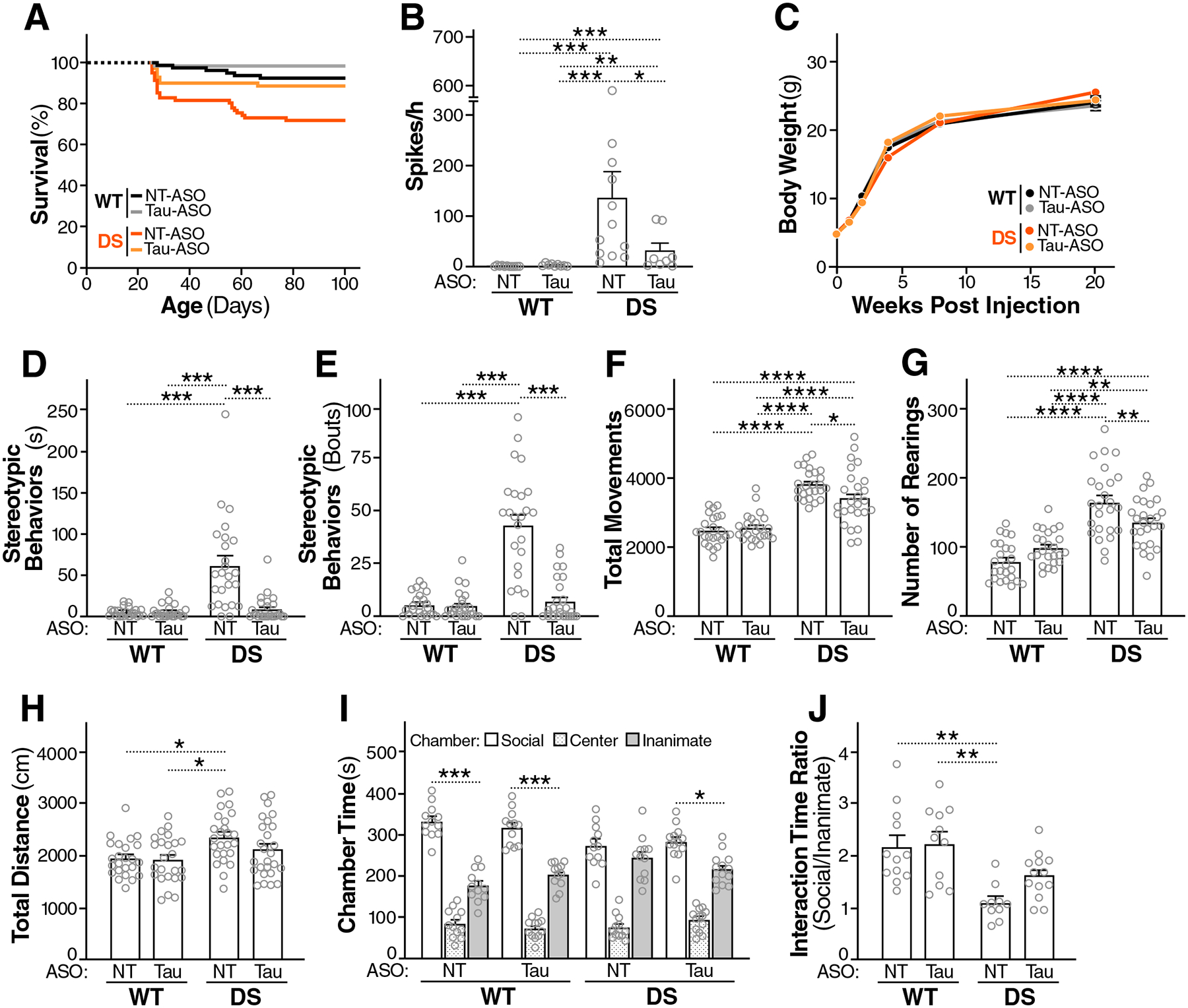
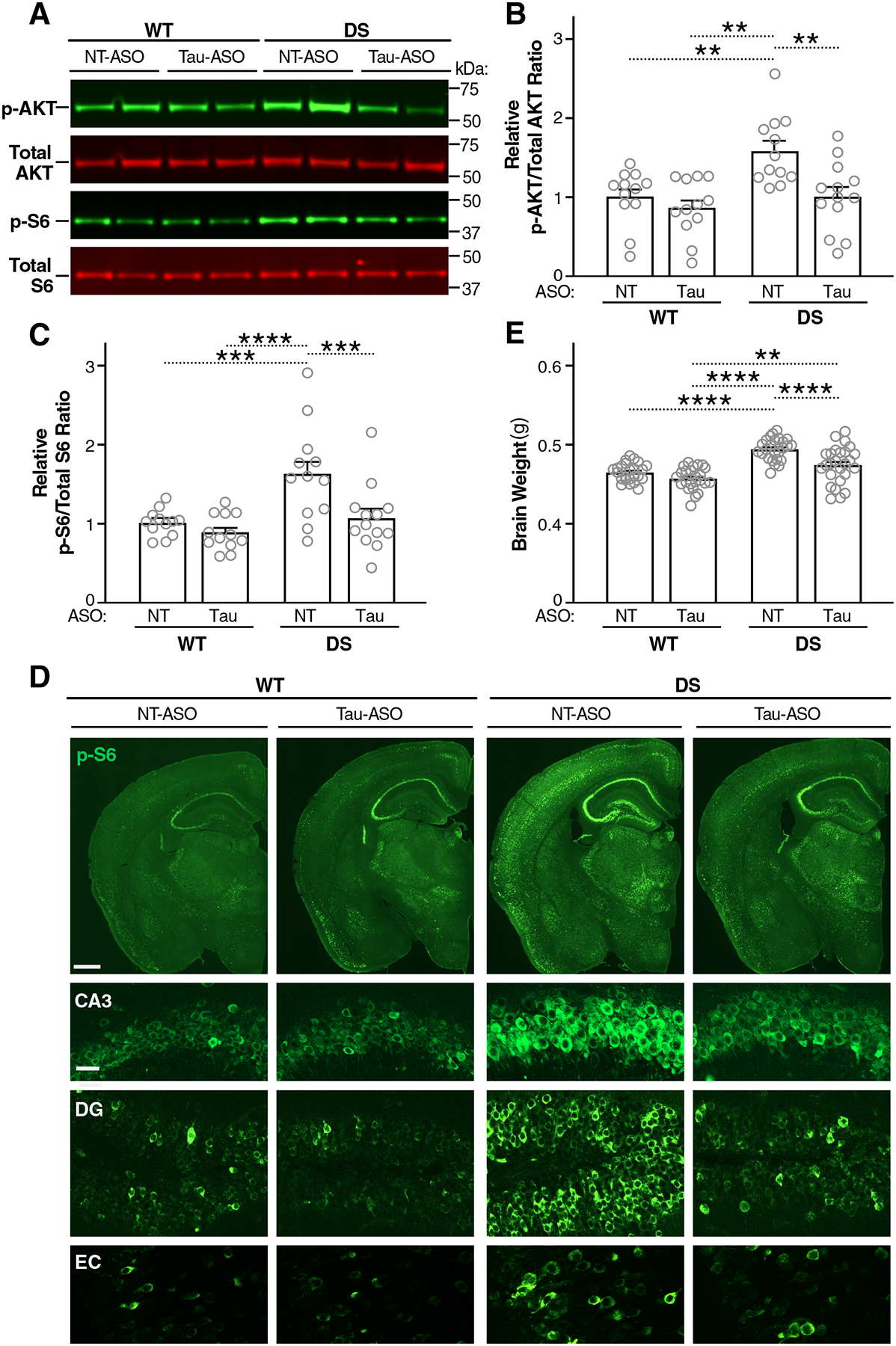
Intracellular accumulation of TAU aggregates is a hallmark of several neurodegenerative diseases. However, global genetic reduction of TAU is beneficial also in models of other brain disorders that lack such TAU pathology, suggesting a pathogenic role of nonaggregated TAU. Here, conditional ablation of TAU in excitatory, but not inhibitory, neurons reduced epilepsy, sudden unexpected death in epilepsy, overactivation of the phosphoinositide 3-kinase-AKT-mammalian target of rapamycin pathway, brain overgrowth (megalencephaly), and autism-like behaviors in a mouse model of Dravet syndrome, a severe epileptic encephalopathy of early childhood. Furthermore, treatment with a TAU-lowering antisense oligonucleotide, initiated on postnatal day 10, had similar therapeutic effects in this mouse model. Our findings suggest that excitatory neurons are the critical cell type in which TAU has to be reduced to counteract brain dysfunctions associated with Dravet syndrome and that overall cerebral TAU reduction could have similar benefits, even when initiated postnatally.
Figures
Comment in
-
One to Rule Them All: A Unique TAU Therapy for Neurodevelopmental Encephalopathies.Epilepsy Curr. 2022 Sep 22;22(6):390-391. doi: 10.1177/15357597221126332. eCollection 2022 Nov-Dec. Epilepsy Curr. 2022. PMID: 36426176 Free PMC article. No abstract available.
References
-
- Fombonne E, MacFarlane H, Salem AC, Epidemiological surveys of ASD: advances and remaining challenges. J. Autism Dev. Disord, Online ahead of print (2021). - PubMed
-
- Satterstrom FK, Kosmicki JA, Wang J, Breen MS, De Rubeis S, An JY, Peng M, Collins R, Grove J, Klei L, Stevens C, Reichert J, Mulhern MS, Artomov M, Gerges S, Sheppard B, Xu X, Bhaduri A, Norman U, Brand H, Schwartz G, Nguyen R, Guerrero EE, Dias C, C. Autism Sequencing, P.-B. C. i, Betancur C, Cook EH, Gallagher L, Gill M, Sutcliffe JS, Thurm A, Zwick ME, Borglum AD, State MW, Cicek AE, Talkowski ME, Cutler DJ, Devlin B, Sanders SJ, Roeder K, Daly MJ, Buxbaum JD, Large-scale exome sequencing study implicates both developmental and functional changes in the neurobiology of autism. Cell 180, 568–584.e23 (2020). - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
