

ARVCF catenin controls force production during vertebrate convergent extension
- PMID: 35476939
- PMCID: PMC9308970
- DOI: 10.1016/j.devcel.2022.04.001
ARVCF catenin controls force production during vertebrate convergent extension
Abstract
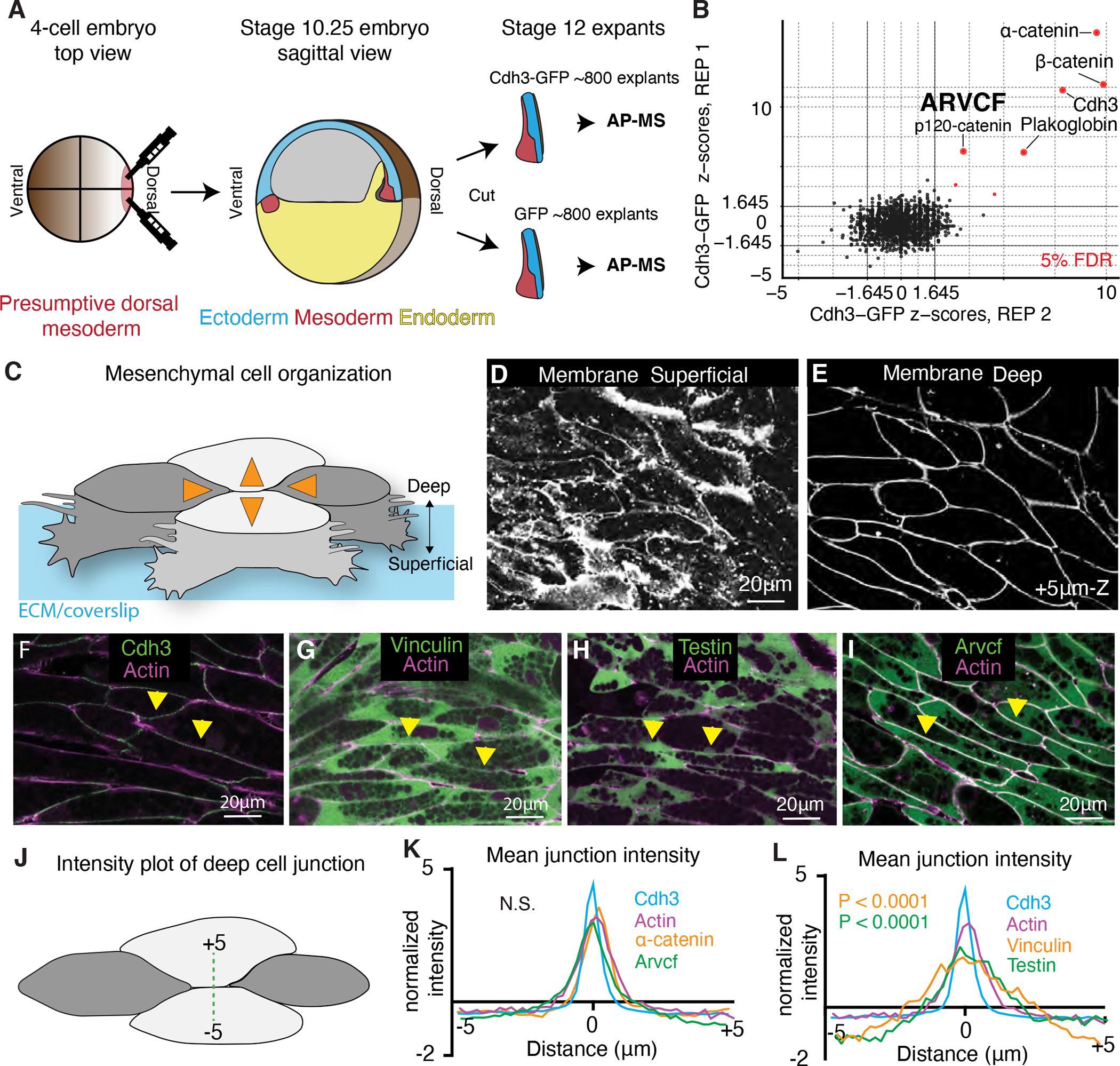
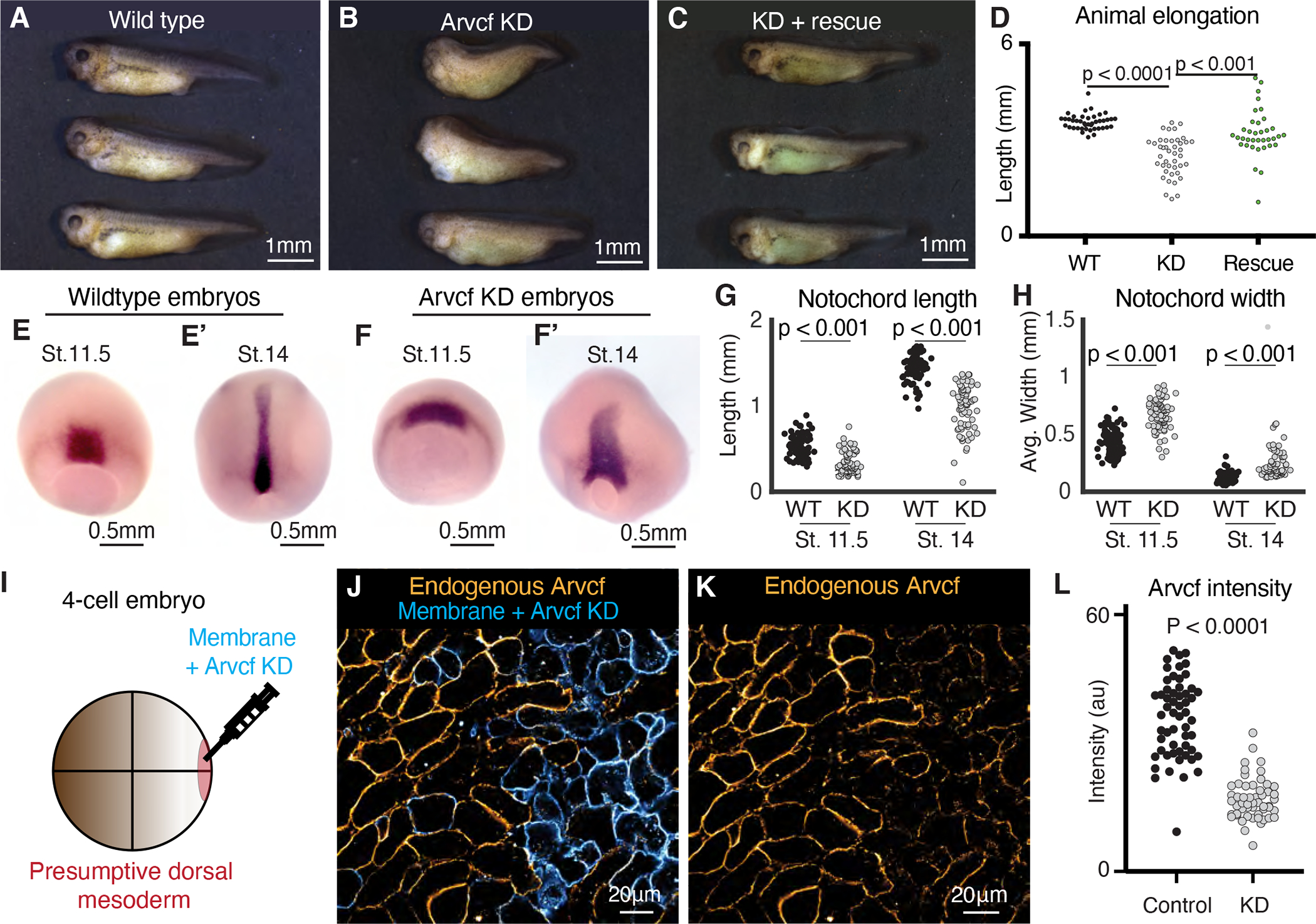
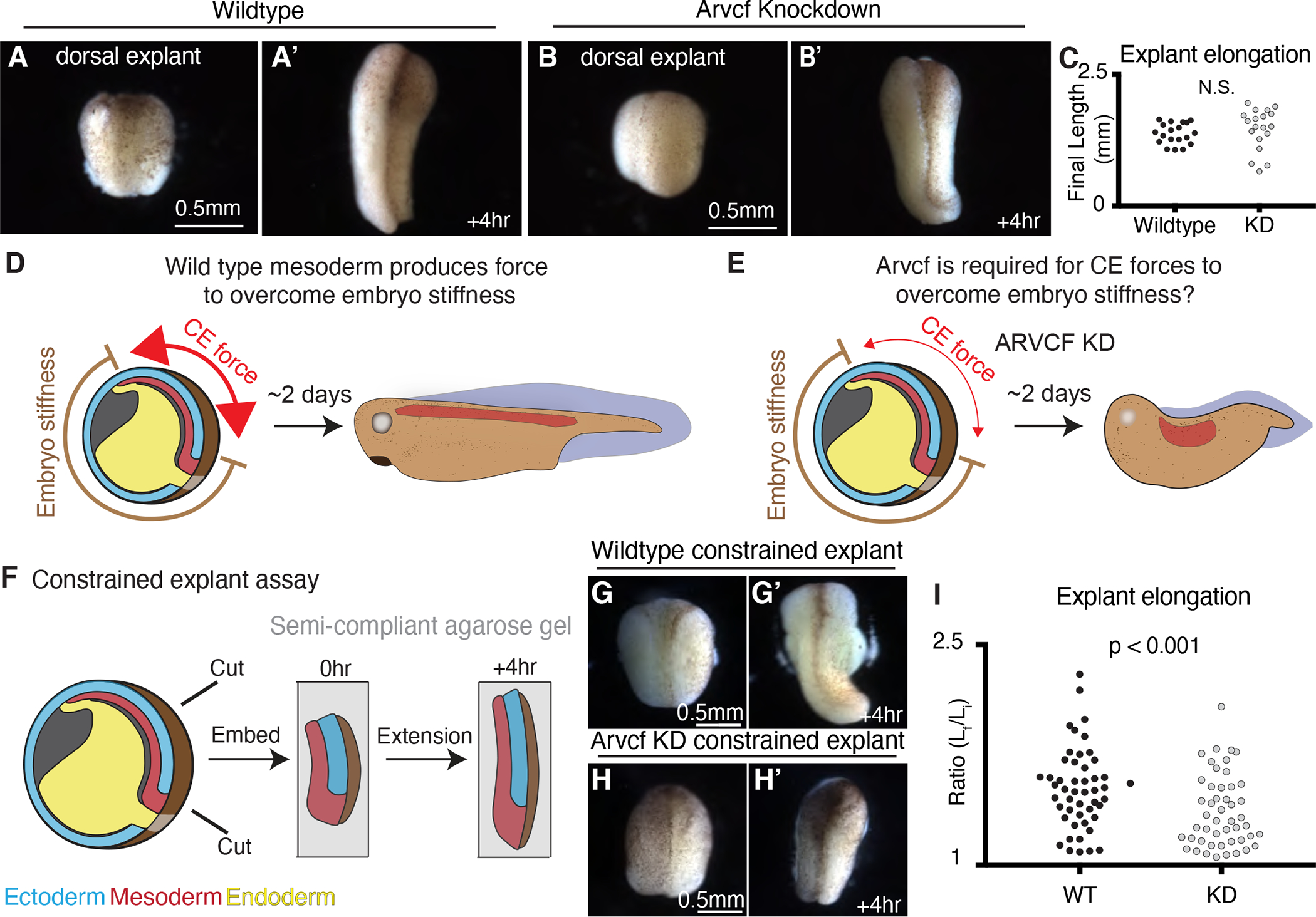
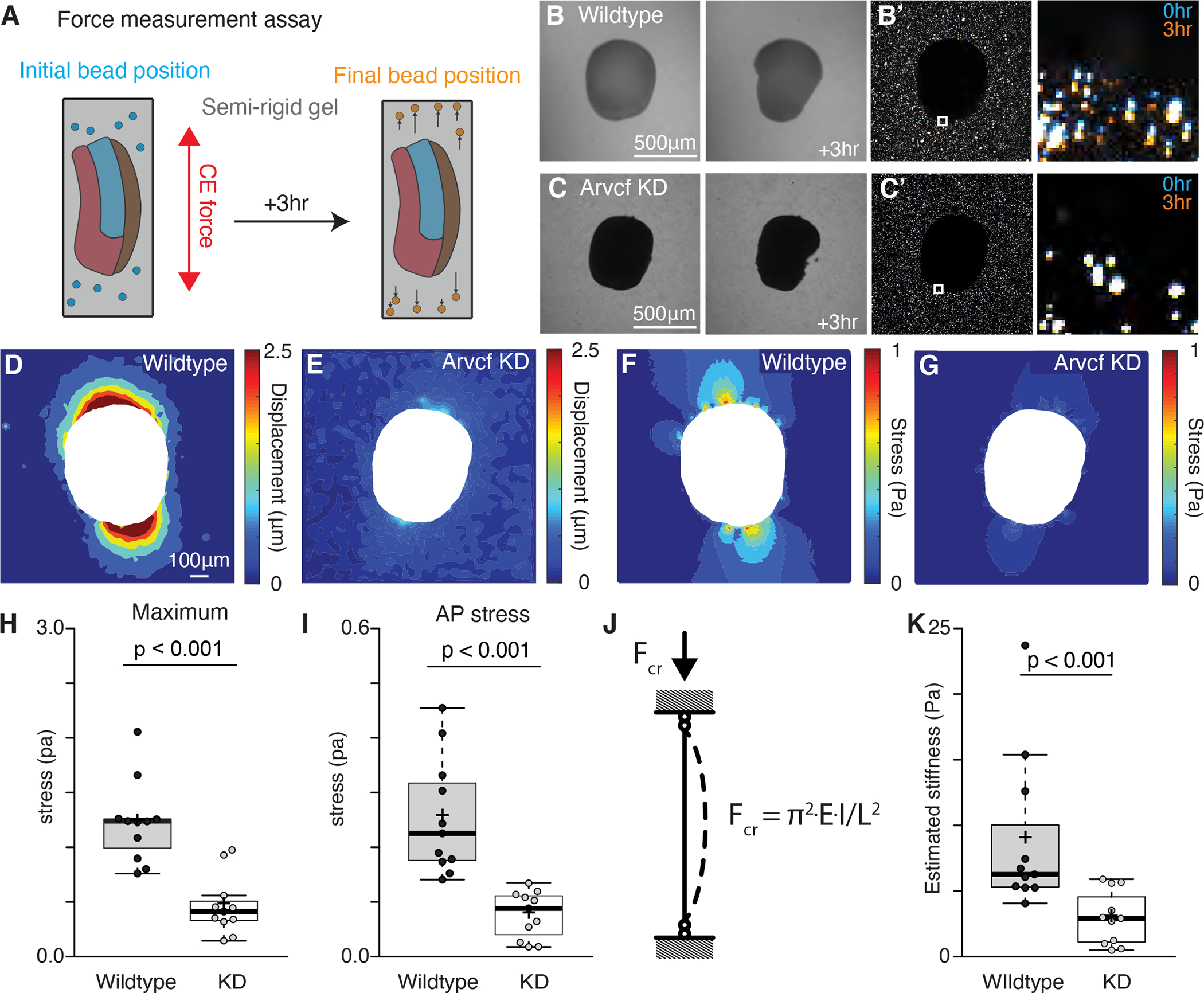
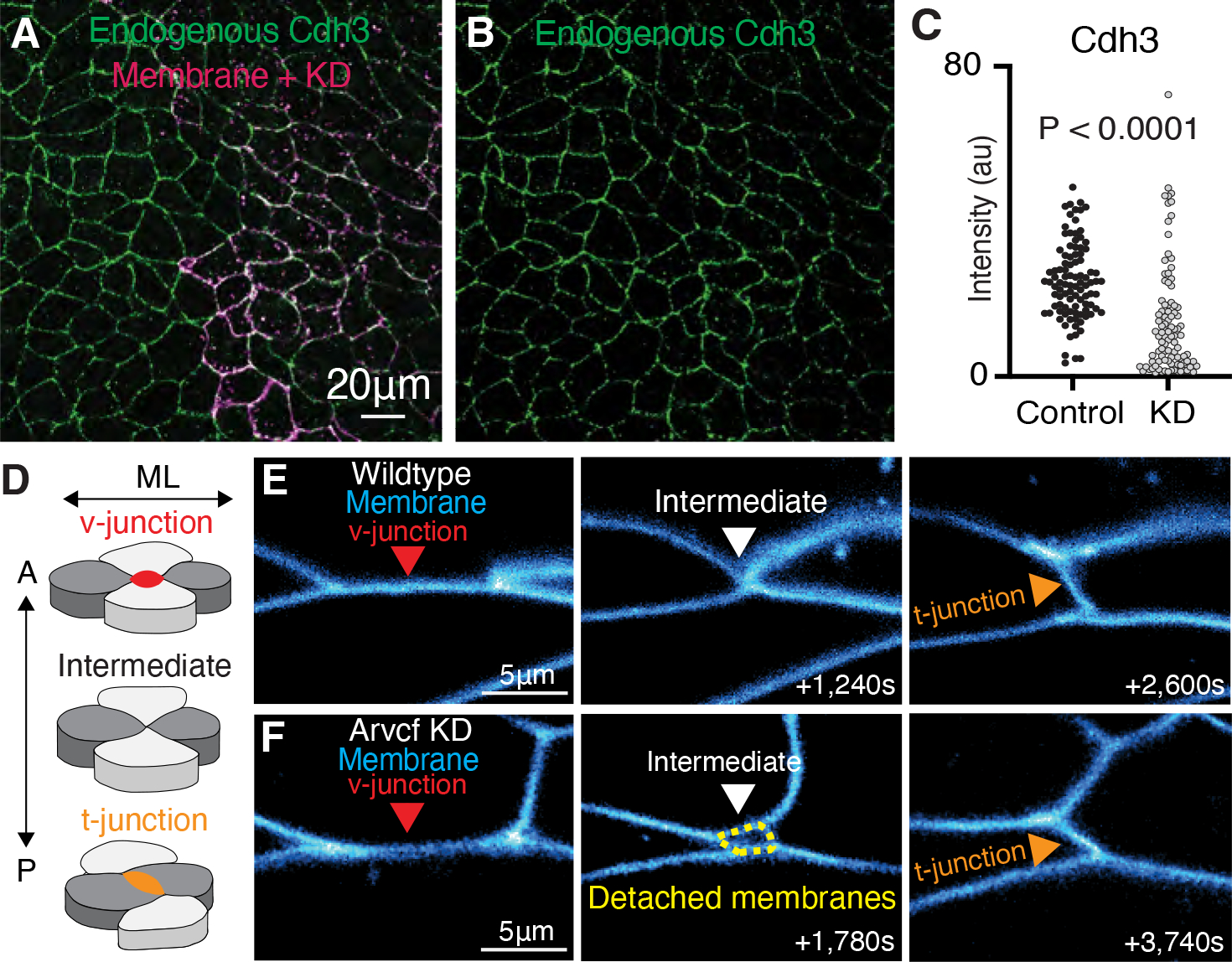
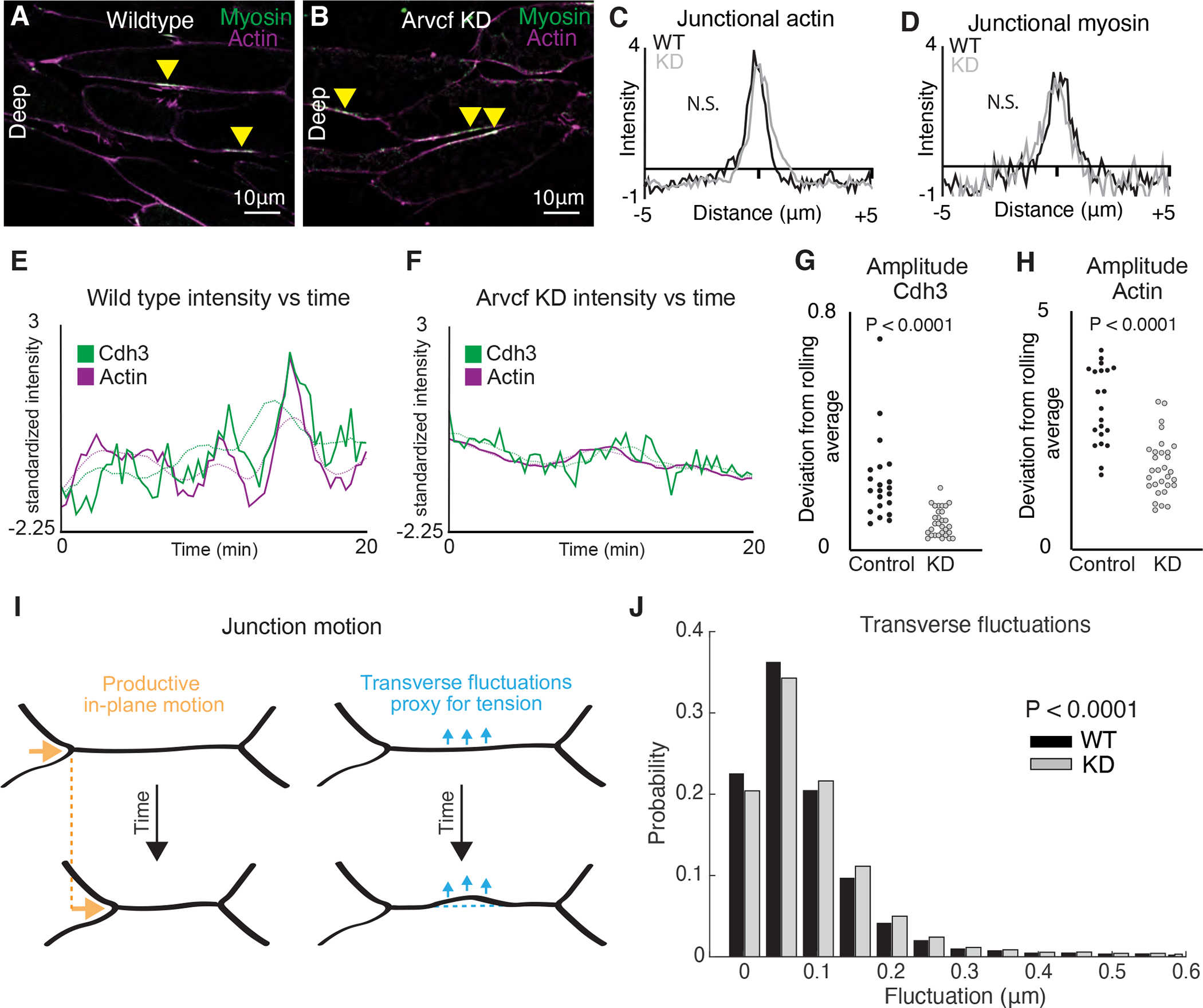
The design of an animal's body plan is encoded in the genome, and the execution of this program is a mechanical progression involving coordinated movement of proteins, cells, and whole tissues. Thus, a challenge to understanding morphogenesis is connecting events that occur across various length scales. Here, we describe how a poorly characterized adhesion effector, Arvcf catenin, controls Xenopus head-to-tail axis extension. We find that Arvcf is required for axis extension within the intact organism but not within isolated tissues. We show that the organism-scale phenotype results from a defect in tissue-scale force production. Finally, we determine that the force defect results from the dampening of the pulsatile recruitment of cell adhesion and cytoskeletal proteins to membranes. These results provide a comprehensive understanding of Arvcf function during axis extension and produce an insight into how a cellular-scale defect in adhesion results in an organism-scale failure of development.
Keywords: Arvcf; biomechanics; cadherin; catenin; cell adhesion; convergent extension; morphogenesis.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.B.W. is a member of the Developmental Cell advisory board.
Figures
References
-
- Badel P, Rohan CP, Avril S, 2013. Finite Element simulation of buckling-induced vein tortuosity and influence of the wall constitutive properties. J Mech Behav Biomed Mater 26, 119–126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
