

GSK3β mediates the spatiotemporal dynamics of NLRP3 inflammasome activation
- PMID: 35477991
- PMCID: PMC9525599
- DOI: 10.1038/s41418-022-00997-y
GSK3β mediates the spatiotemporal dynamics of NLRP3 inflammasome activation
Abstract
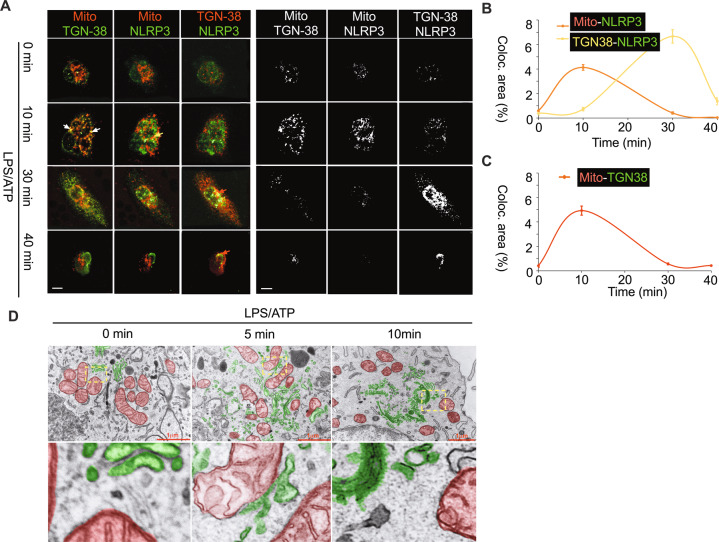
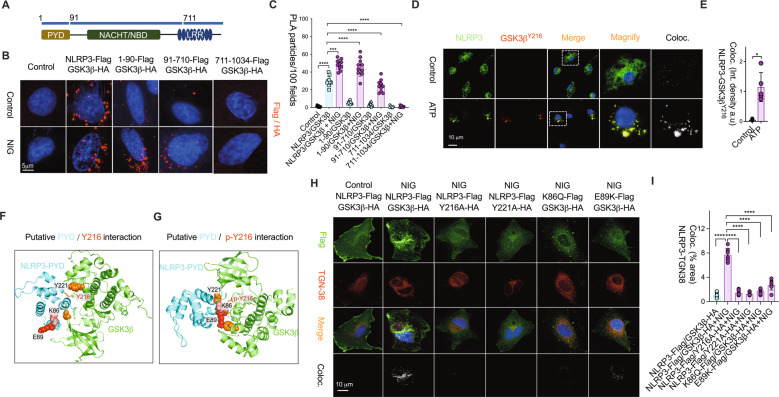
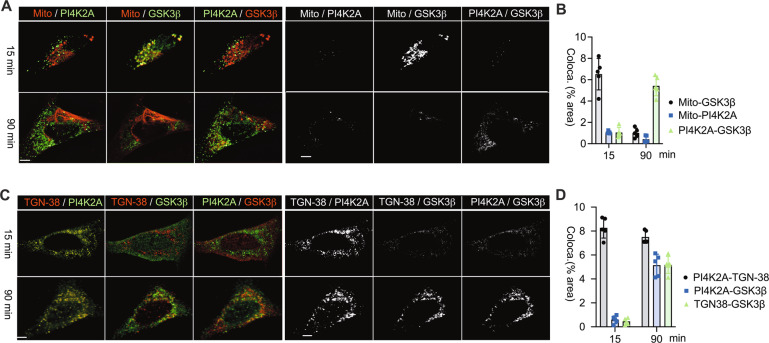
Subcellular machinery of NLRP3 is essential for inflammasome assembly and activation. However, the stepwise process and mechanistic basis of NLRP3 engagement with organelles remain unclear. Herein, we demonstrated glycogen synthase kinase 3β (GSK3β) as a molecular determinant for the spatiotemporal dynamics of NLRP3 inflammasome activation. Using live cell multispectral time-lapse tracking acquisition, we observed that upon stimuli NLRP3 was transiently associated with mitochondria and subsequently recruited to the Golgi network (TGN) where it was retained for inflammasome assembly. This occurred in relation to the temporal contact of mitochondria to Golgi apparatus. NLRP3 stimuli initiate GSK3β activation with subsequent binding to NLRP3, facilitating NLRP3 recruitment to mitochondria and transition to TGN. GSK3β activation also phosphorylates phosphatidylinositol 4-kinase 2 Α (PI4k2A) in TGN to promote sustained NLRP3 oligomerization. Our study has identified the interplay between GSK3β signaling and the organelles dynamics of NLRP3 required for inflammasome activation and opens new avenues for therapeutic intervention.
© 2022. The Author(s), under exclusive licence to ADMC Associazione Differenziamento e Morte Cellulare.
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
