

Structural characterisation of amyloid-like fibrils formed by an amyloidogenic peptide segment of β-lactoglobulin
- PMID: 35480736
- PMCID: PMC9037834
- DOI: 10.1039/d1ra03575d
Structural characterisation of amyloid-like fibrils formed by an amyloidogenic peptide segment of β-lactoglobulin
Abstract
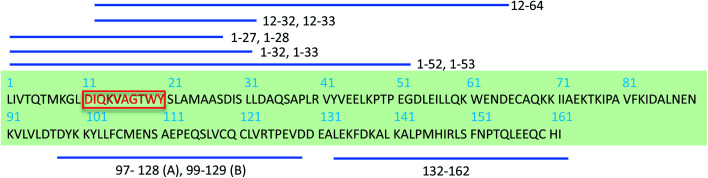
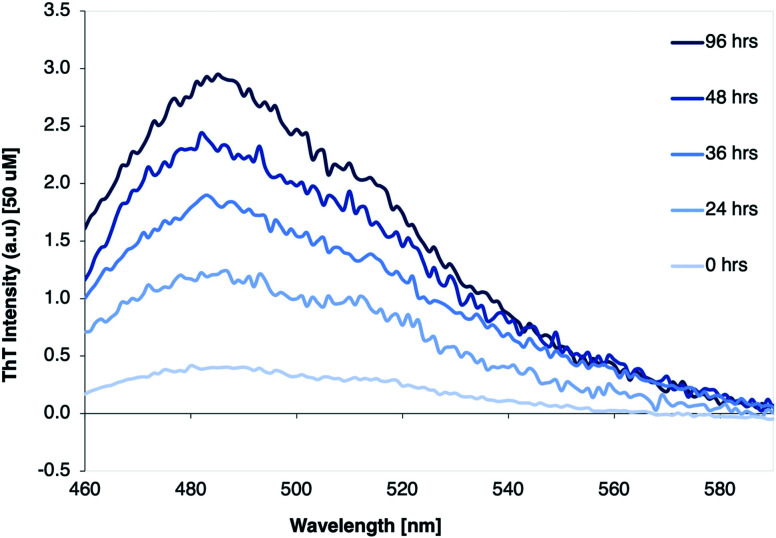
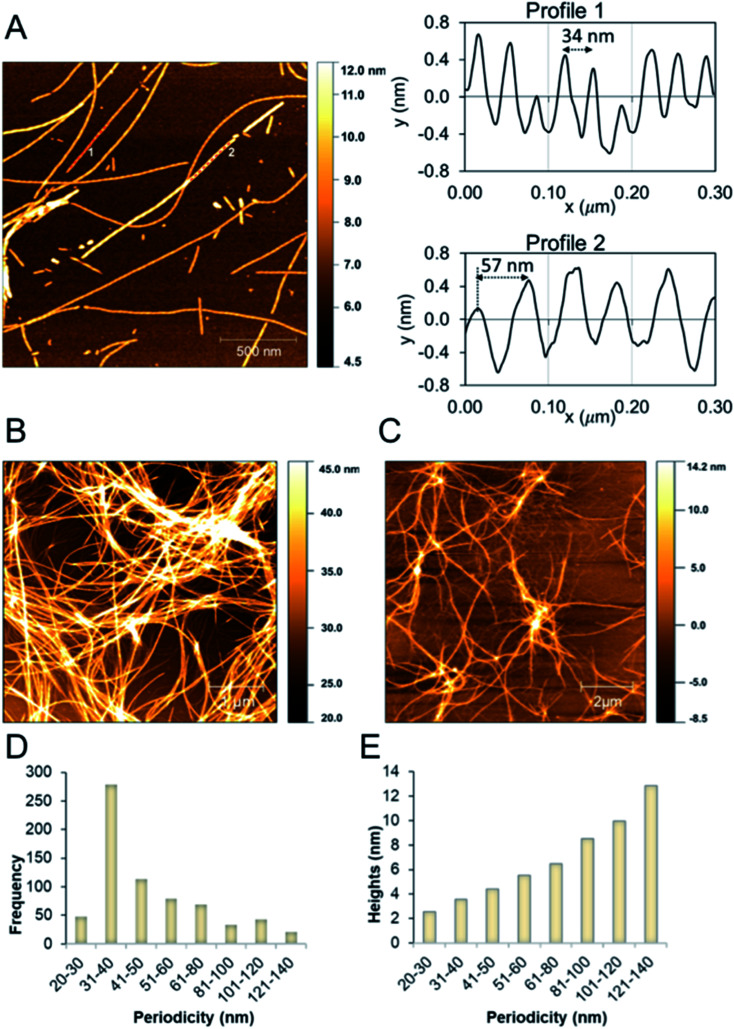
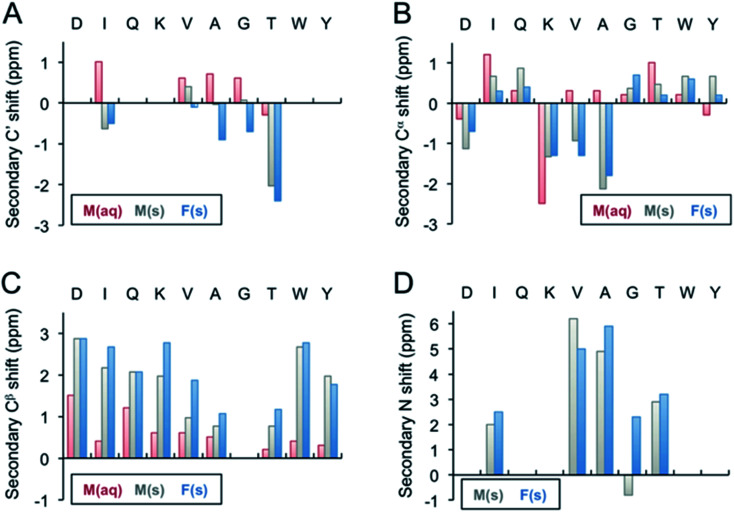
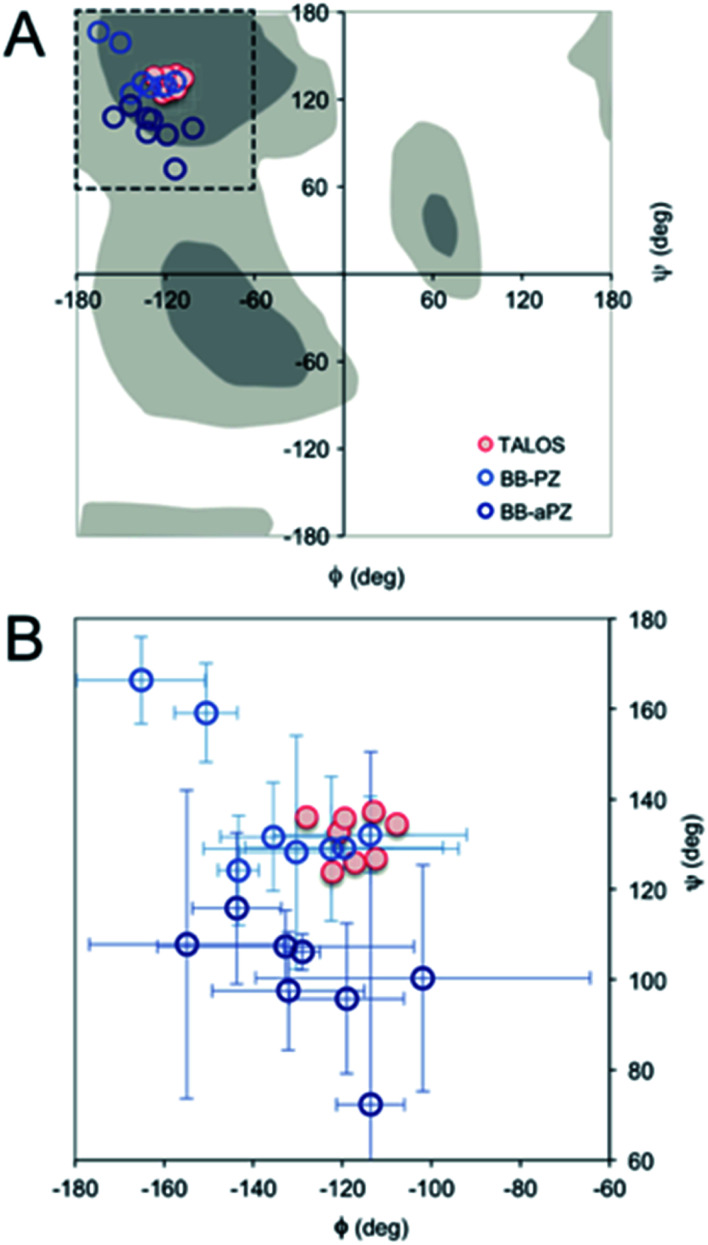
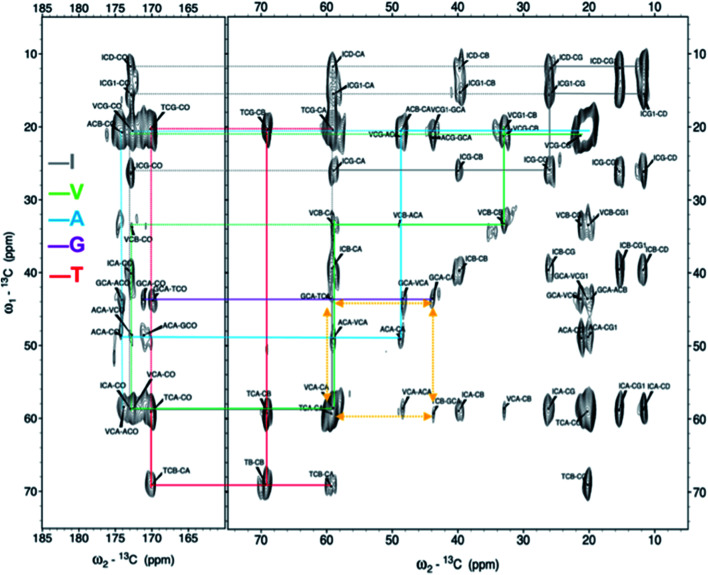
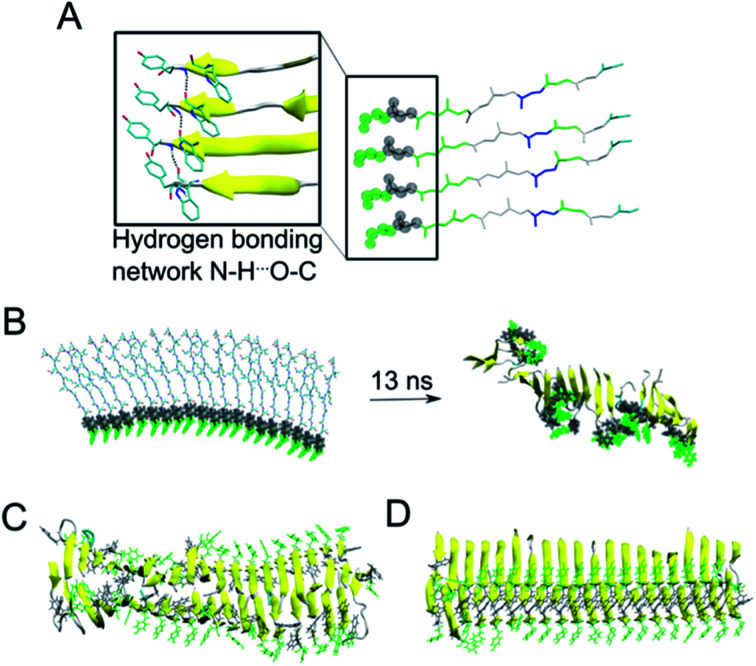
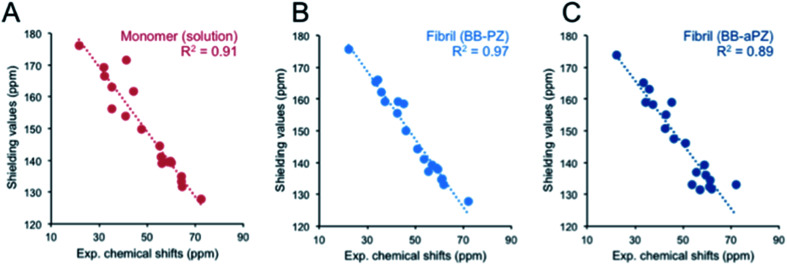
Protein nanofibrils (PNFs) represent a promising class of biobased nanomaterials for biomedical and materials science applications. In the design of such materials, a fundamental understanding of the structure-function relationship at both molecular and nanoscale levels is essential. Here we report investigations of the nanoscale morphology and molecular arrangement of amyloid-like PNFs of a synthetic peptide fragment consisting of residues 11-20 of the protein β-lactoglobulin (β-LG11-20), an important model system for PNF materials. Nanoscale fibril morphology was analysed by atomic force microscopy (AFM) that indicates the presence of polymorphic self-assembly of protofilaments. However, observation of a single set of 13C and 15N resonances in the solid-state NMR spectra for the β-LG11-20 fibrils suggests that the observed polymorphism originates from the assembly of protofilaments at the nanoscale but not from the molecular structure. The secondary structure and inter-residue proximities in the β-LG11-20 fibrils were probed using NMR experiments of the peptide with 13C- and 15N-labelled amino acid residues at selected positions. We can conclude that the peptides form parallel β-sheets, but the NMR data was inconclusive regarding inter-sheet packing. Molecular dynamics simulations confirm the stability of parallel β-sheets and suggest two preferred modes of packing. Comparison of molecular dynamics models with NMR data and calculated chemical shifts indicates that both packing models are possible.
This journal is © The Royal Society of Chemistry.
Conflict of interest statement
There are no conflicts to declare.
Figures
Similar articles
-
Supramolecular structural constraints on Alzheimer's beta-amyloid fibrils from electron microscopy and solid-state nuclear magnetic resonance.Biochemistry. 2002 Dec 24;41(51):15436-50. doi: 10.1021/bi0204185. Biochemistry. 2002. PMID: 12484785
-
Amyloid fibril formation by A beta 16-22, a seven-residue fragment of the Alzheimer's beta-amyloid peptide, and structural characterization by solid state NMR.Biochemistry. 2000 Nov 14;39(45):13748-59. doi: 10.1021/bi0011330. Biochemistry. 2000. PMID: 11076514
-
Design of parallel 𝛽-sheet nanofibrils using Monte Carlo search, coarse-grained simulations, and experimental testing.Protein Sci. 2024 Aug;33(8):e5102. doi: 10.1002/pro.5102. Protein Sci. 2024. PMID: 39037281 Free PMC article.
-
Solid-state NMR as a method to reveal structure and membrane-interaction of amyloidogenic proteins and peptides.Biochim Biophys Acta. 2007 Aug;1768(8):1900-12. doi: 10.1016/j.bbamem.2007.03.025. Epub 2007 Apr 5. Biochim Biophys Acta. 2007. PMID: 17524351 Review.
-
[Molecular mechanism of amyloid formation by Ab peptide: review of own works].Biomed Khim. 2018 Jan;64(1):94-109. doi: 10.18097/PBMC20186401094. Biomed Khim. 2018. PMID: 29460839 Review. Russian.
Cited by
-
Mechanism of action of IC 100, a humanized IgG4 monoclonal antibody targeting apoptosis-associated speck-like protein containing a caspase recruitment domain (ASC).Transl Res. 2023 Jan;251:27-40. doi: 10.1016/j.trsl.2022.06.016. Epub 2022 Jul 3. Transl Res. 2023. PMID: 35793783 Free PMC article.
-
Food protein-derived amyloids do not accelerate amyloid β aggregation.Sci Rep. 2023 Jan 31;13(1):985. doi: 10.1038/s41598-023-28147-5. Sci Rep. 2023. PMID: 36720893 Free PMC article.
-
Biodegradable and Implantable Triboelectric Nanogenerator Improved by β-Lactoglobulin Fibrils-Assisted Flexible PVA Porous Film.Adv Sci (Weinh). 2025 Jun;12(24):e2409914. doi: 10.1002/advs.202409914. Epub 2024 Nov 11. Adv Sci (Weinh). 2025. PMID: 39526831 Free PMC article.
-
Protein nanofibrils and their use as building blocks of sustainable materials.RSC Adv. 2021 Dec 8;11(62):39188-39215. doi: 10.1039/d1ra06878d. eCollection 2021 Dec 6. RSC Adv. 2021. PMID: 35492452 Free PMC article. Review.
-
Ultrasound-induced protein restructuring and ordered aggregation to form amyloid crystals.Eur Biophys J. 2022 Jul;51(4-5):335-352. doi: 10.1007/s00249-022-01601-4. Epub 2022 May 16. Eur Biophys J. 2022. PMID: 35576075 Free PMC article.
References
-
- V. N. Uversky and A. L. Fink, Protein Misfolding, Aggregation, and Conformational Diseases: Part B: Molecular Mechanisms of Conformational Diseases, Springer US, Boston, MA, 2007, pp. 61–110
LinkOut - more resources
Full Text Sources
Miscellaneous