

Biological noise is a key determinant of the reproducibility and adaptability of cardiac pacemaking and EC coupling
- PMID: 35482009
- PMCID: PMC9059386
- DOI: 10.1085/jgp.202012613
Biological noise is a key determinant of the reproducibility and adaptability of cardiac pacemaking and EC coupling
Abstract
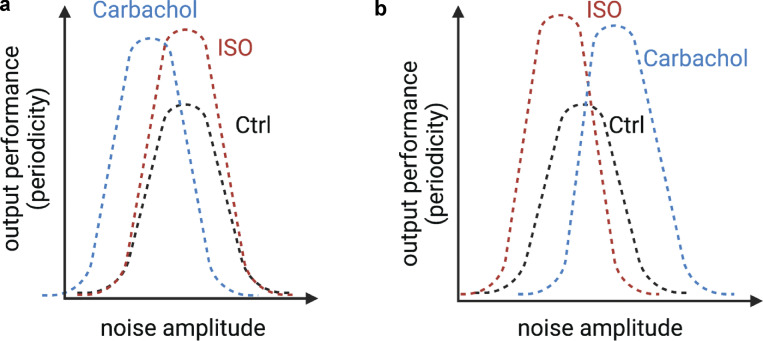
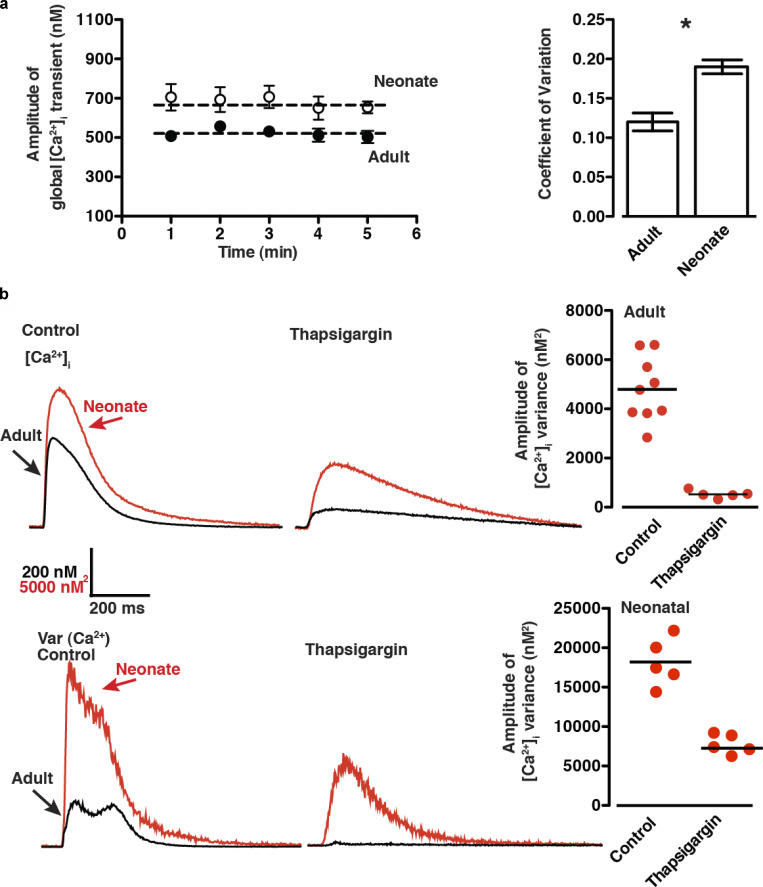
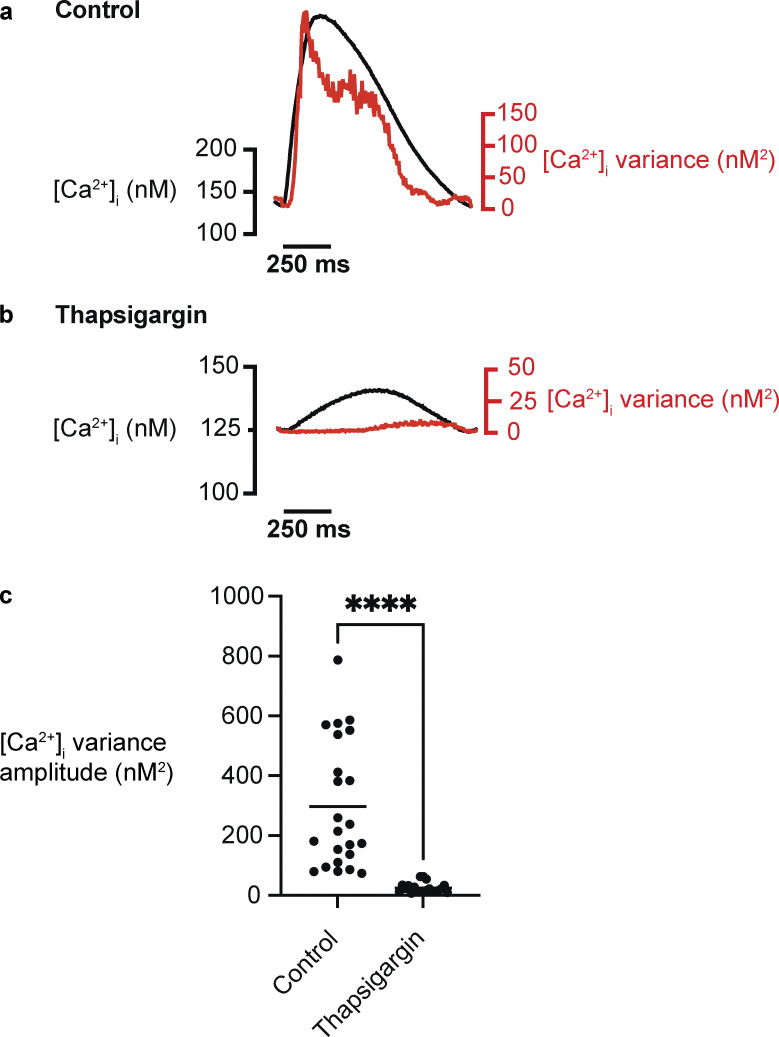
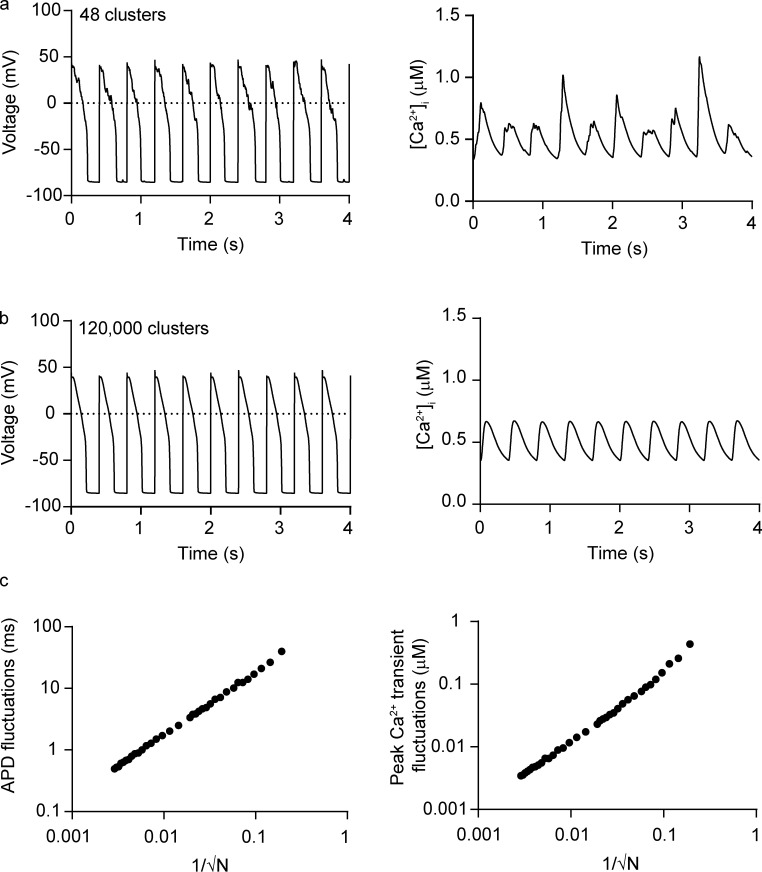
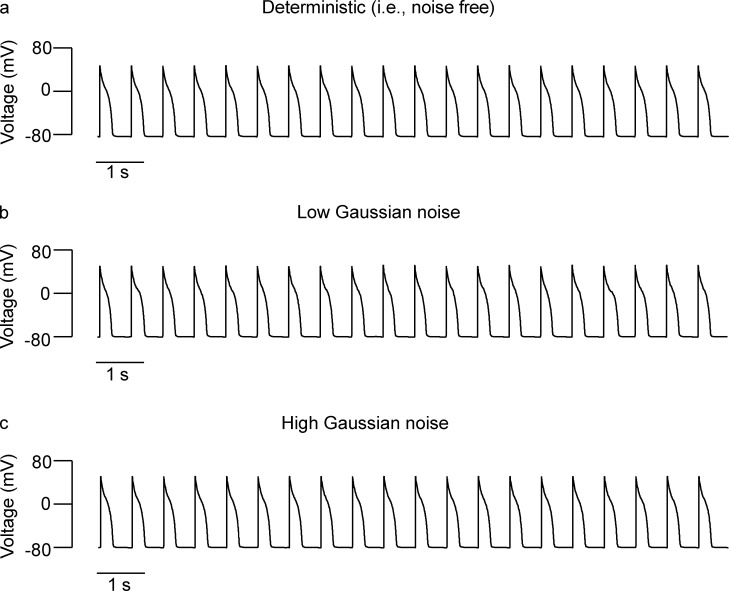
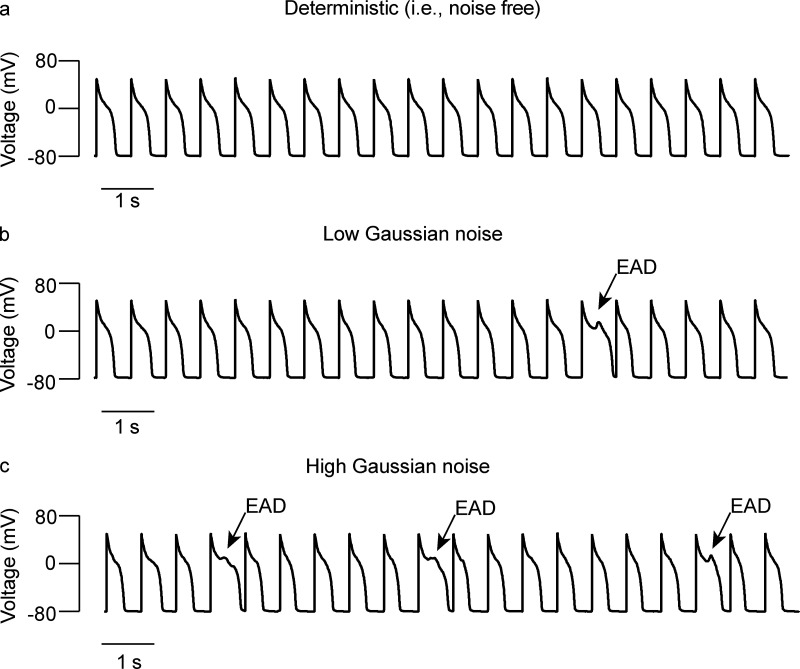
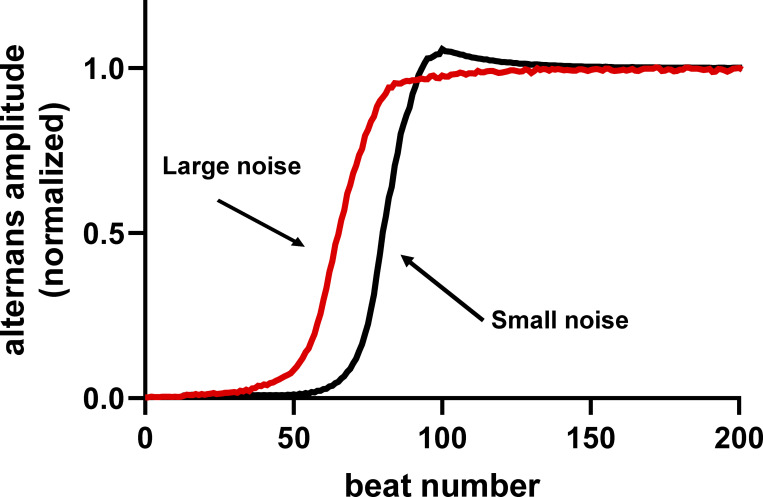
Each heartbeat begins with the generation of an action potential in pacemaking cells in the sinoatrial node. This signal triggers contraction of cardiac muscle through a process termed excitation-contraction (EC) coupling. EC coupling is initiated in dyadic structures of cardiac myocytes, where ryanodine receptors in the junctional sarcoplasmic reticulum come into close apposition with clusters of CaV1.2 channels in invaginations of the sarcolemma. Cooperative activation of CaV1.2 channels within these clusters causes a local increase in intracellular Ca2+ that activates the juxtaposed ryanodine receptors. A salient feature of healthy cardiac function is the reliable and precise beat-to-beat pacemaking and amplitude of Ca2+ transients during EC coupling. In this review, we discuss recent discoveries suggesting that the exquisite reproducibility of this system emerges, paradoxically, from high variability at subcellular, cellular, and network levels. This variability is attributable to stochastic fluctuations in ion channel trafficking, clustering, and gating, as well as dyadic structure, which increase intracellular Ca2+ variance during EC coupling. Although the effects of these large, local fluctuations in function and organization are sometimes negligible at the macroscopic level owing to spatial-temporal summation within and across cells in the tissue, recent work suggests that the "noisiness" of these intracellular Ca2+ events may either enhance or counterintuitively reduce variability in a context-dependent manner. Indeed, these noisy events may represent distinct regulatory features in the tuning of cardiac contractility. Collectively, these observations support the importance of incorporating experimentally determined values of Ca2+ variance in all EC coupling models. The high reproducibility of cardiac contraction is a paradoxical outcome of high Ca2+ signaling variability at subcellular, cellular, and network levels caused by stochastic fluctuations in multiple processes in time and space. This underlying stochasticity, which counterintuitively manifests as reliable, consistent Ca2+ transients during EC coupling, also allows for rapid changes in cardiac rhythmicity and contractility in health and disease.
© 2022 Guarina et al.
Figures
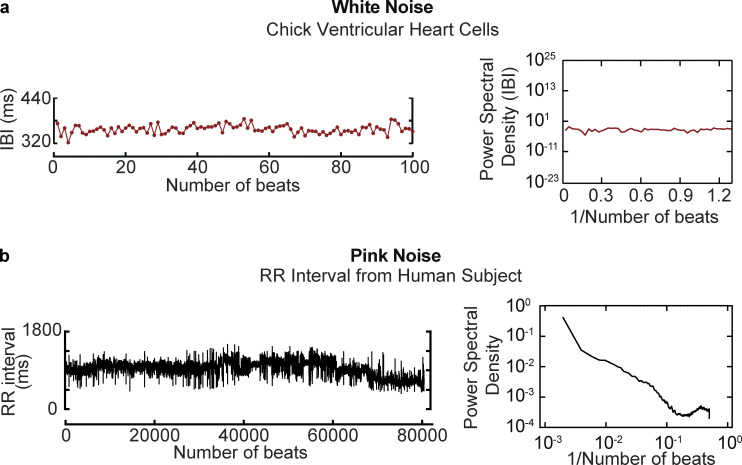
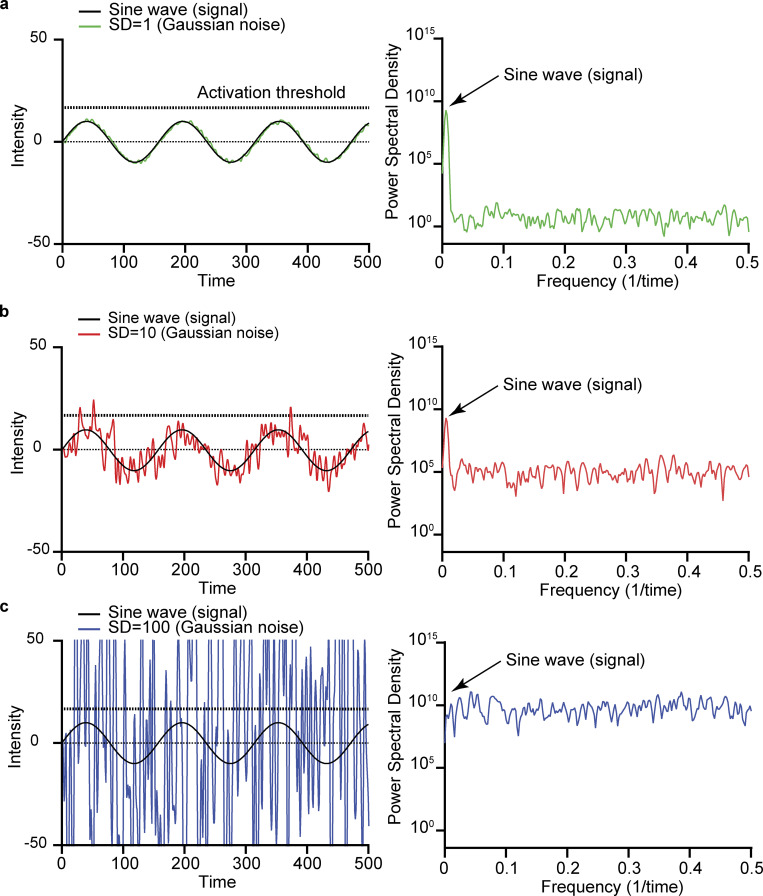
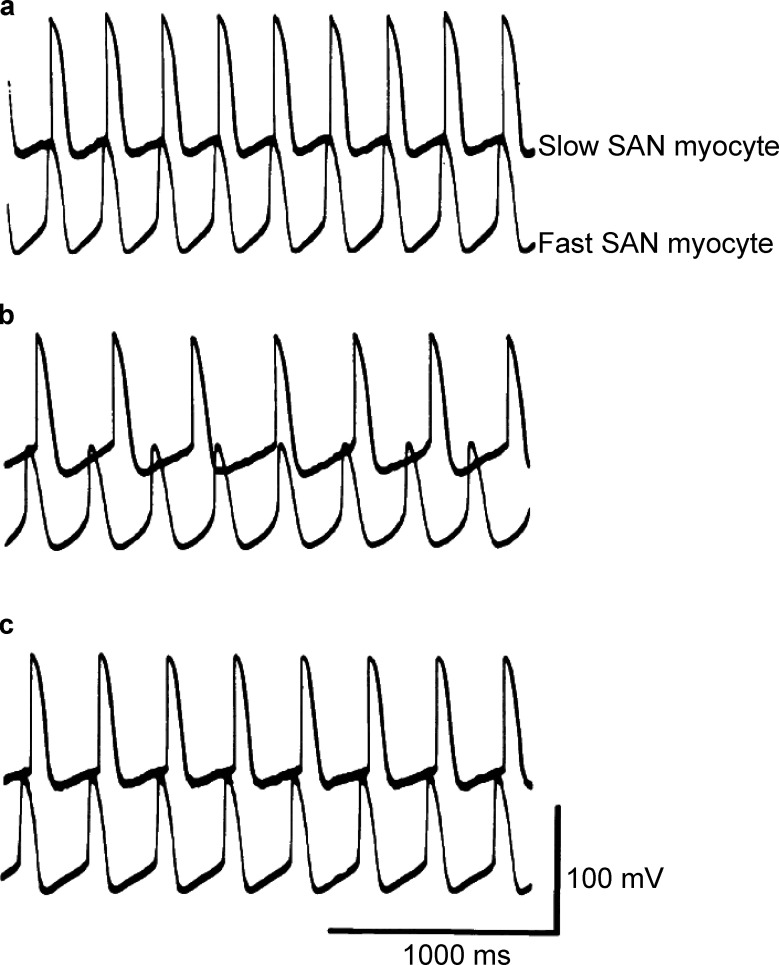
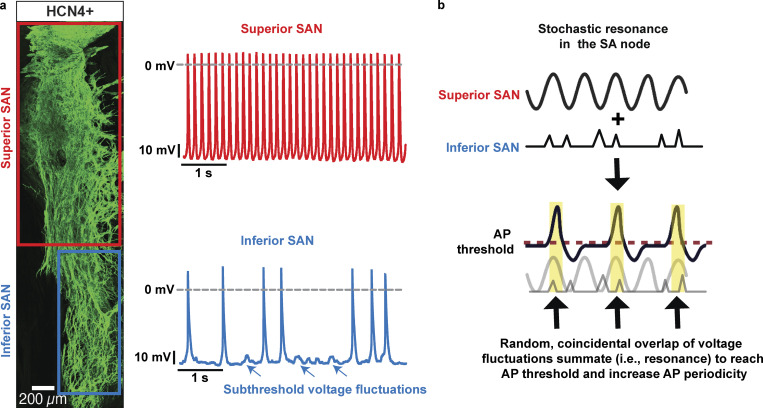
Similar articles
-
Ca2+ Channels Mediate Bidirectional Signaling between Sarcolemma and Sarcoplasmic Reticulum in Muscle Cells.Cells. 2019 Dec 24;9(1):55. doi: 10.3390/cells9010055. Cells. 2019. PMID: 31878335 Free PMC article. Review.
-
Nanoscale organization of cardiac calcium channels is dependent on thyroid hormone status.Am J Physiol Heart Circ Physiol. 2024 Nov 1;327(5):H1309-H1326. doi: 10.1152/ajpheart.00272.2024. Epub 2024 Oct 4. Am J Physiol Heart Circ Physiol. 2024. PMID: 39365674 Free PMC article.
-
A novel mechanism of tandem activation of ryanodine receptors by cytosolic and SR luminal Ca2+ during excitation-contraction coupling in atrial myocytes.J Physiol. 2017 Jun 15;595(12):3835-3845. doi: 10.1113/JP273611. Epub 2017 Feb 1. J Physiol. 2017. PMID: 28028837 Free PMC article.
-
Ryanodine receptor sensitivity governs the stability and synchrony of local calcium release during cardiac excitation-contraction coupling.J Mol Cell Cardiol. 2016 Mar;92:82-92. doi: 10.1016/j.yjmcc.2016.01.024. Epub 2016 Jan 28. J Mol Cell Cardiol. 2016. PMID: 26827896 Free PMC article.
-
Calcium signaling between sarcolemmal calcium channels and ryanodine receptors in heart cells.Front Biosci. 2002 Sep 1;7:d1867-78. doi: 10.2741/A885. Front Biosci. 2002. PMID: 12161336 Review.
Cited by
-
The Central Brain of the Heart: The Sinoatrial Node.JACC Clin Electrophysiol. 2022 Oct;8(10):1216-1218. doi: 10.1016/j.jacep.2022.08.016. JACC Clin Electrophysiol. 2022. PMID: 36265996 Free PMC article. No abstract available.
-
Adenosine reduces sinoatrial node cell action potential firing rate by uncoupling its membrane and calcium clocks.Front Physiol. 2022 Nov 24;13:977807. doi: 10.3389/fphys.2022.977807. eCollection 2022. Front Physiol. 2022. PMID: 36505046 Free PMC article.
-
A computational model predicts sex-specific responses to calcium channel blockers in mammalian mesenteric vascular smooth muscle.bioRxiv [Preprint]. 2024 Jan 6:2023.06.24.546394. doi: 10.1101/2023.06.24.546394. bioRxiv. 2024. Update in: Elife. 2024 Feb 09;12:RP90604. doi: 10.7554/eLife.90604. PMID: 37425682 Free PMC article. Updated. Preprint.
-
Disorder in Ca2+ release unit locations confers robustness but cuts flexibility of heart pacemaking.J Gen Physiol. 2022 Sep 5;154(9):e202113061. doi: 10.1085/jgp.202113061. Epub 2022 Aug 9. J Gen Physiol. 2022. PMID: 35943725 Free PMC article.
-
The formation of KV2.1 macro-clusters is required for sex-specific differences in L-type CaV1.2 clustering and function in arterial myocytes.Res Sq [Preprint]. 2023 Jul 19:rs.3.rs-3136085. doi: 10.21203/rs.3.rs-3136085/v1. Res Sq. 2023. Update in: Commun Biol. 2023 Nov 14;6(1):1165. doi: 10.1038/s42003-023-05527-1. PMID: 37502980 Free PMC article. Updated. Preprint.
References
-
- Angelini, M., Pezhouman A., Savalli N., Chang M.G., Steccanella F., Scranton K., Calmettes G., Ottolia M., Pantazis A., Karagueuzian H.S., et al. . 2021. Suppression of ventricular arrhythmias by targeting late L-type Ca2+ current. J. Gen. Physiol. 153:e202012584. 10.1085/jgp.202012584 - DOI - PMC - PubMed
