

Ancestry-inclusive dog genomics challenges popular breed stereotypes
- PMID: 35482869
- PMCID: PMC9675396
- DOI: 10.1126/science.abk0639
Ancestry-inclusive dog genomics challenges popular breed stereotypes
Abstract
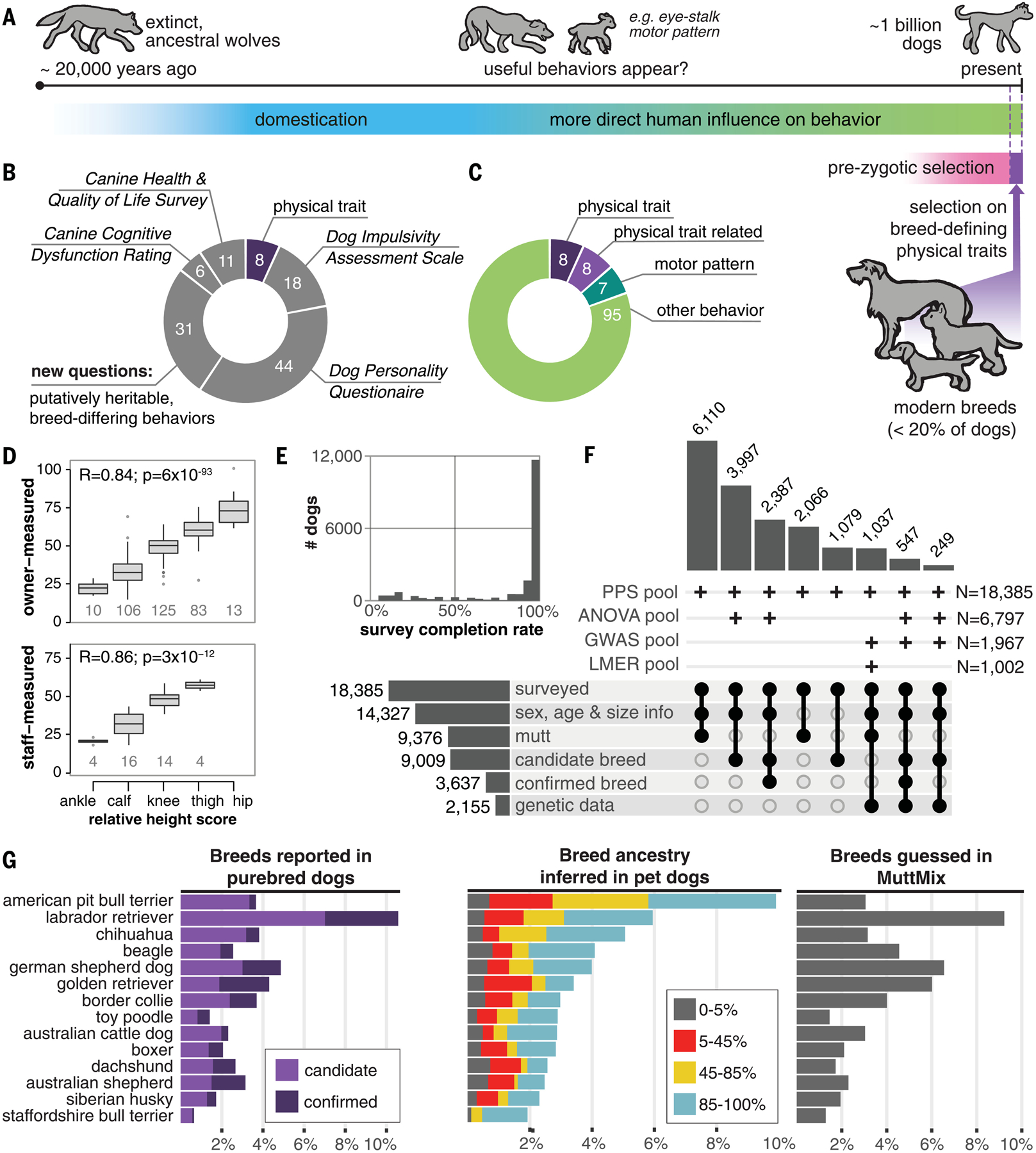
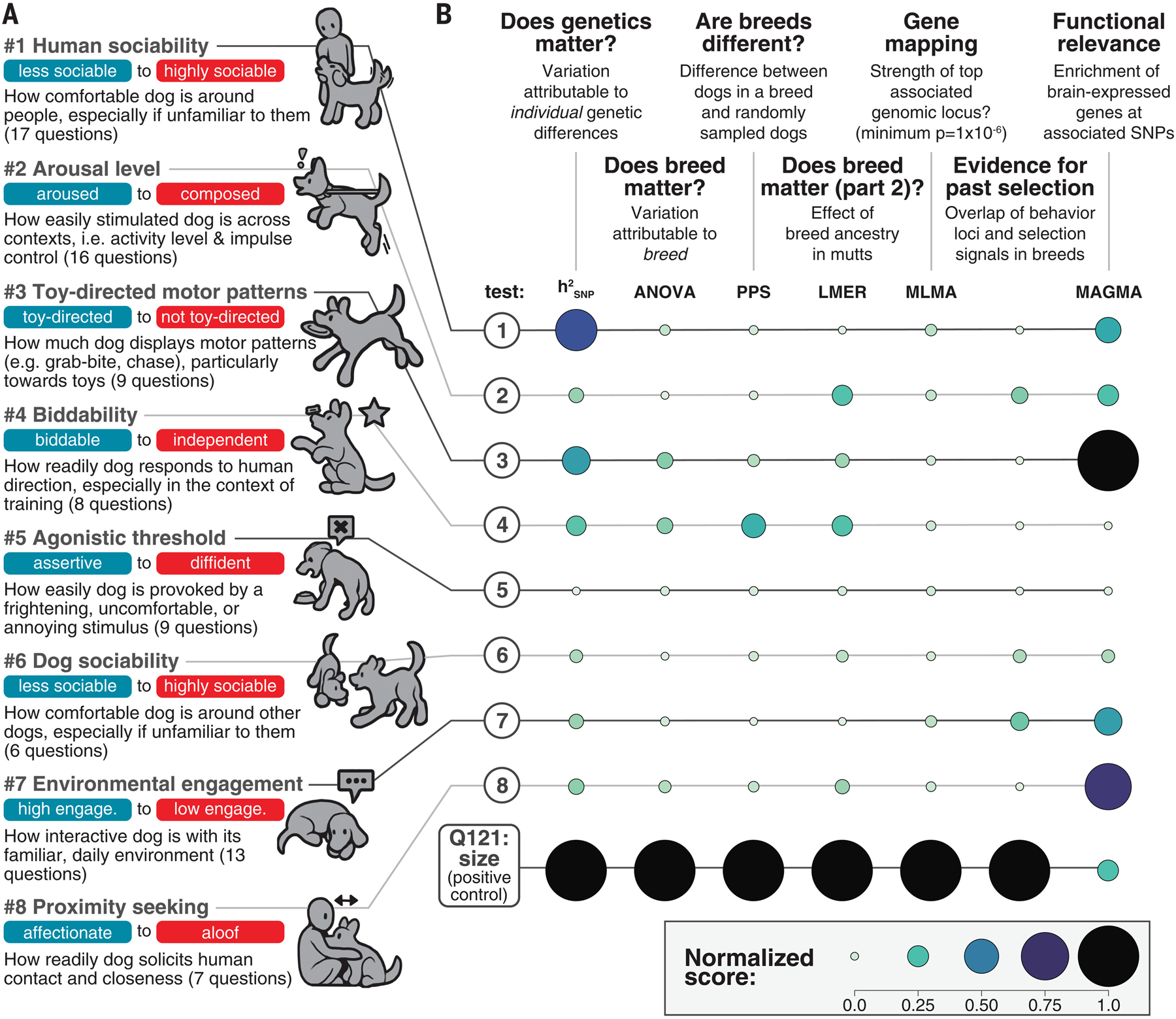
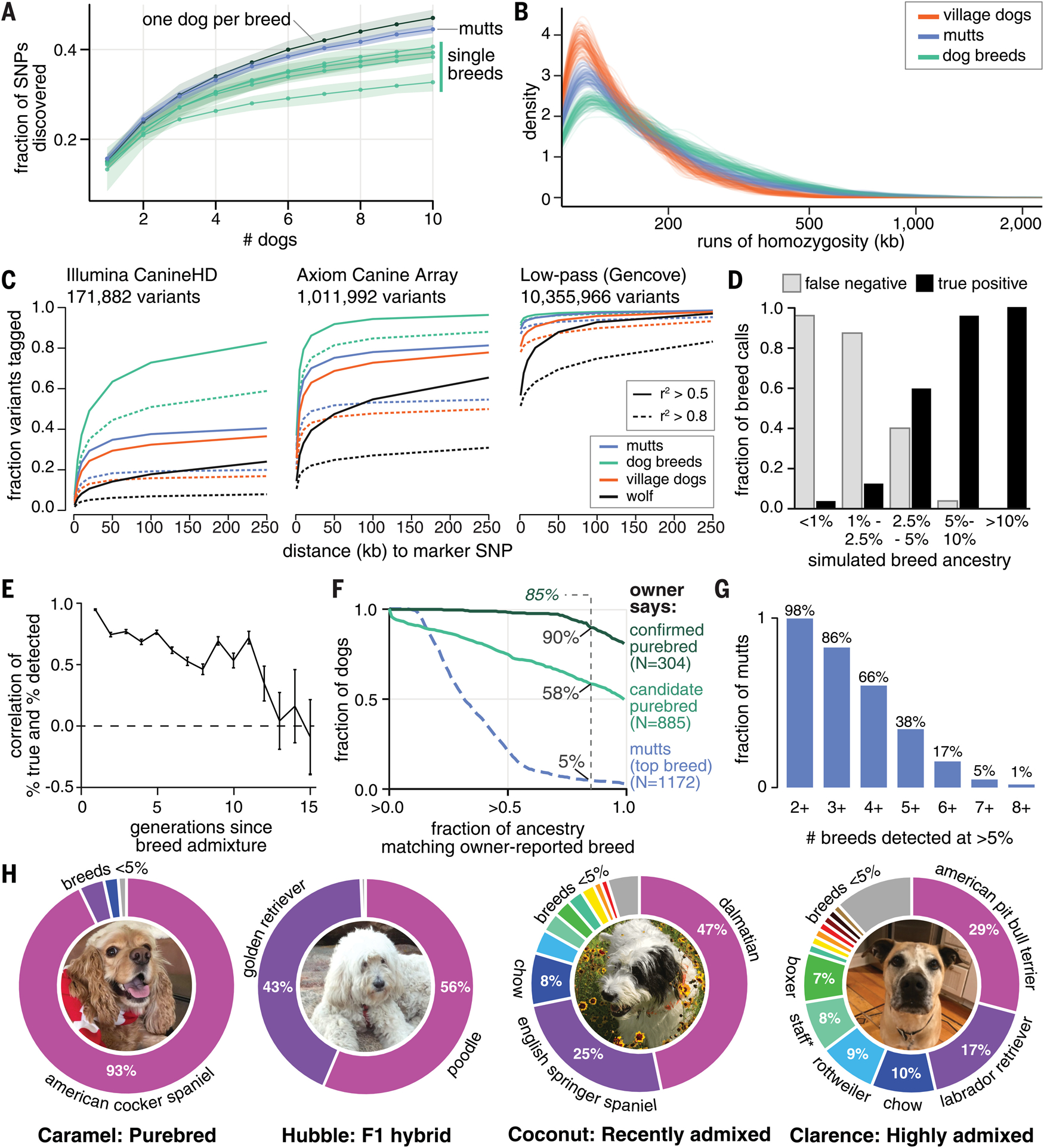
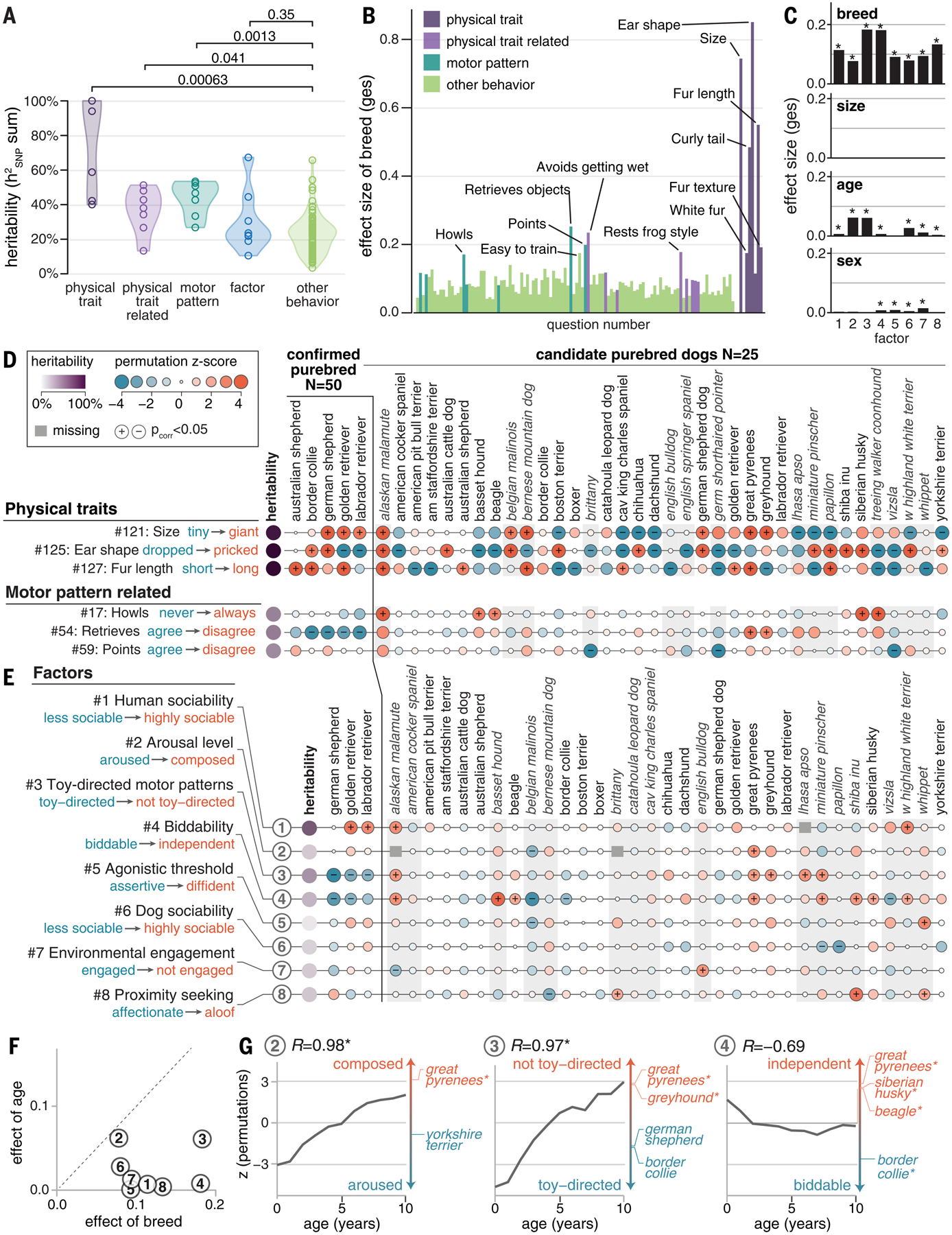
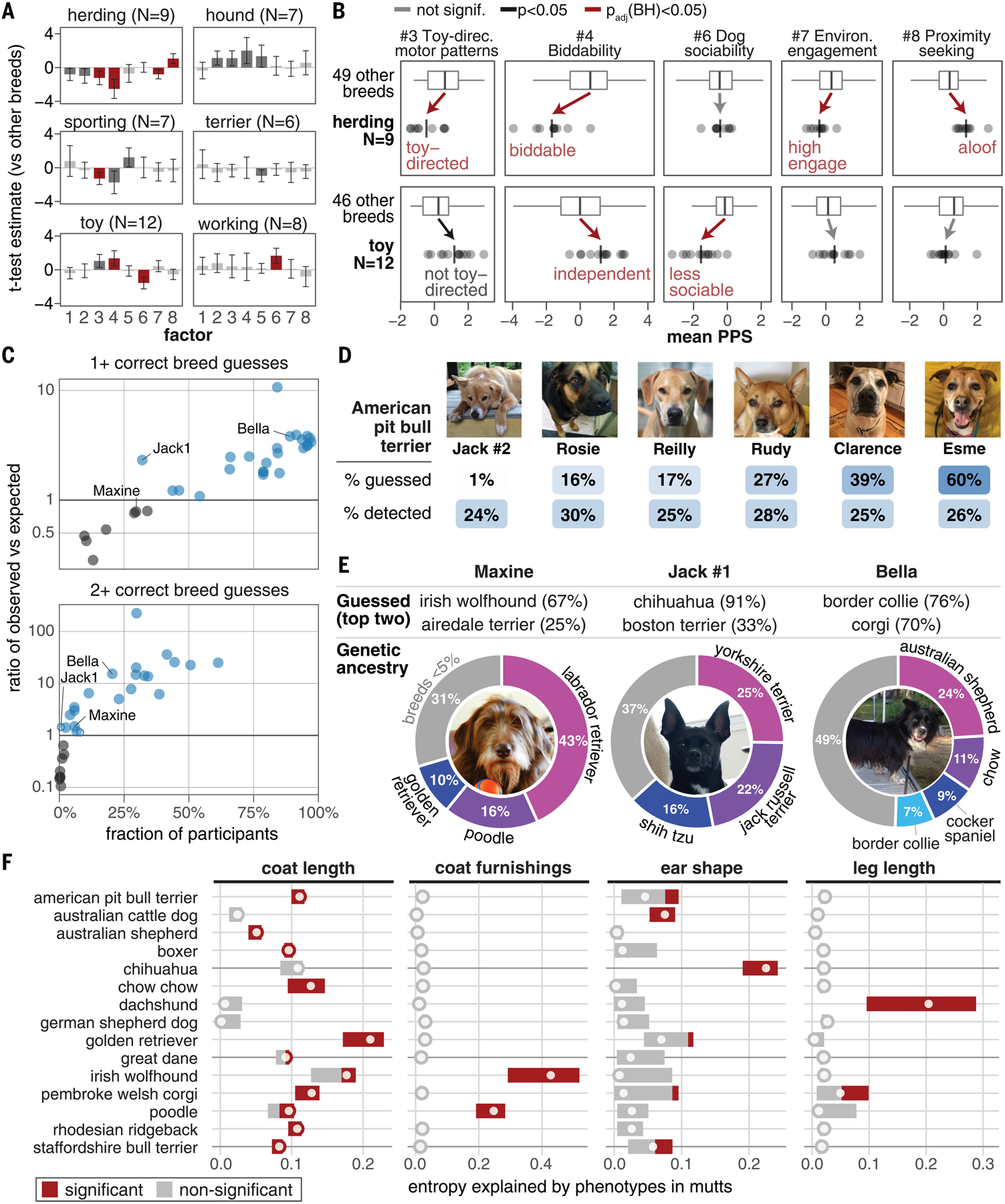
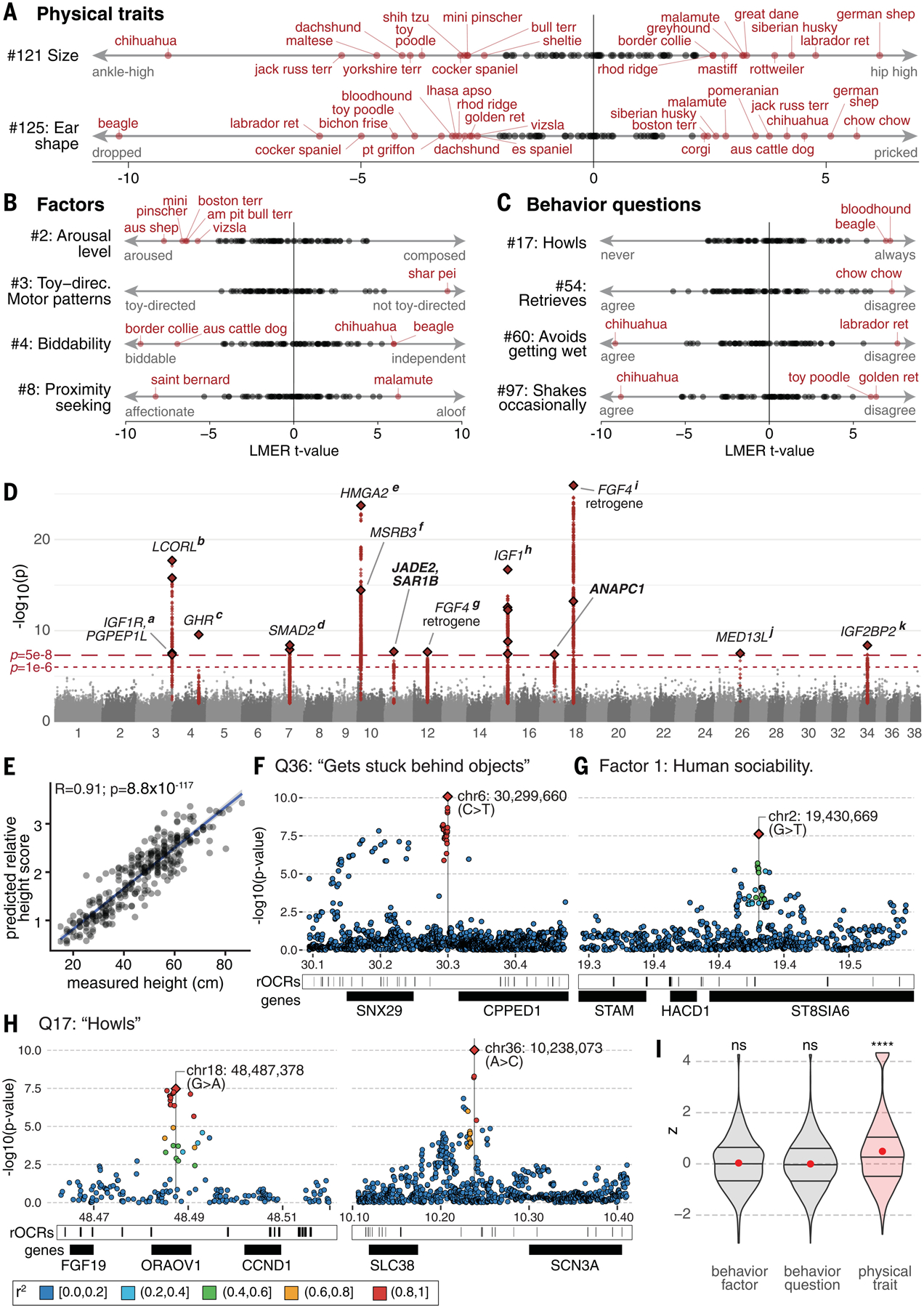
Behavioral genetics in dogs has focused on modern breeds, which are isolated subgroups with distinctive physical and, purportedly, behavioral characteristics. We interrogated breed stereotypes by surveying owners of 18,385 purebred and mixed-breed dogs and genotyping 2155 dogs. Most behavioral traits are heritable [heritability (h2) > 25%], and admixture patterns in mixed-breed dogs reveal breed propensities. Breed explains just 9% of behavioral variation in individuals. Genome-wide association analyses identify 11 loci that are significantly associated with behavior, and characteristic breed behaviors exhibit genetic complexity. Behavioral loci are not unusually differentiated in breeds, but breed propensities align, albeit weakly, with ancestral function. We propose that behaviors perceived as characteristic of modern breeds derive from thousands of years of polygenic adaptation that predates breed formation, with modern breeds distinguished primarily by aesthetic traits.
Conflict of interest statement
Figures
References
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
