

Chronic inflammatory arthritis drives systemic changes in circadian energy metabolism
- PMID: 35482925
- PMCID: PMC9170023
- DOI: 10.1073/pnas.2112781119
Chronic inflammatory arthritis drives systemic changes in circadian energy metabolism
Abstract
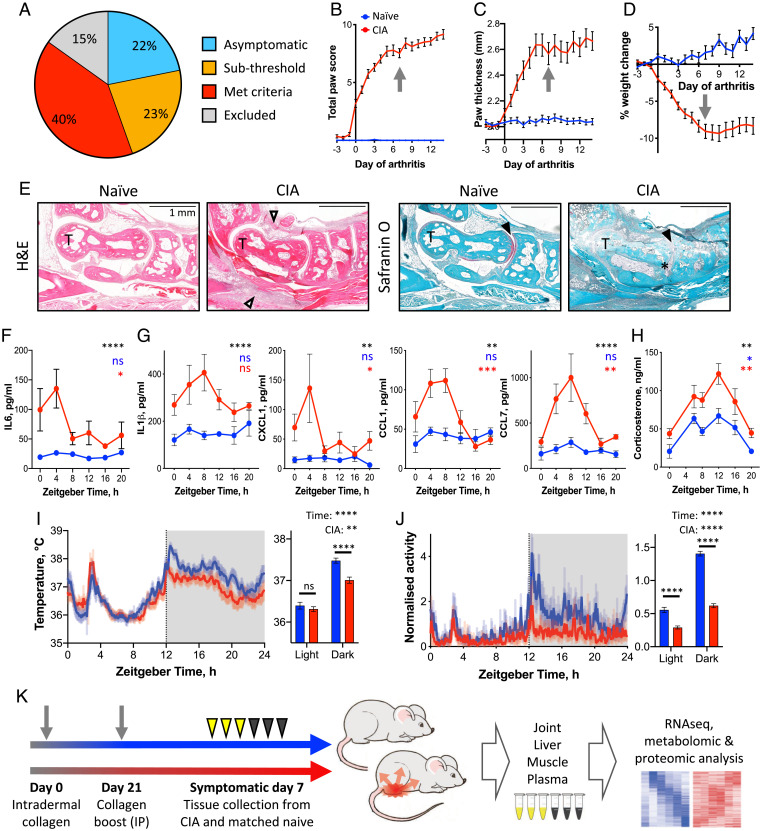
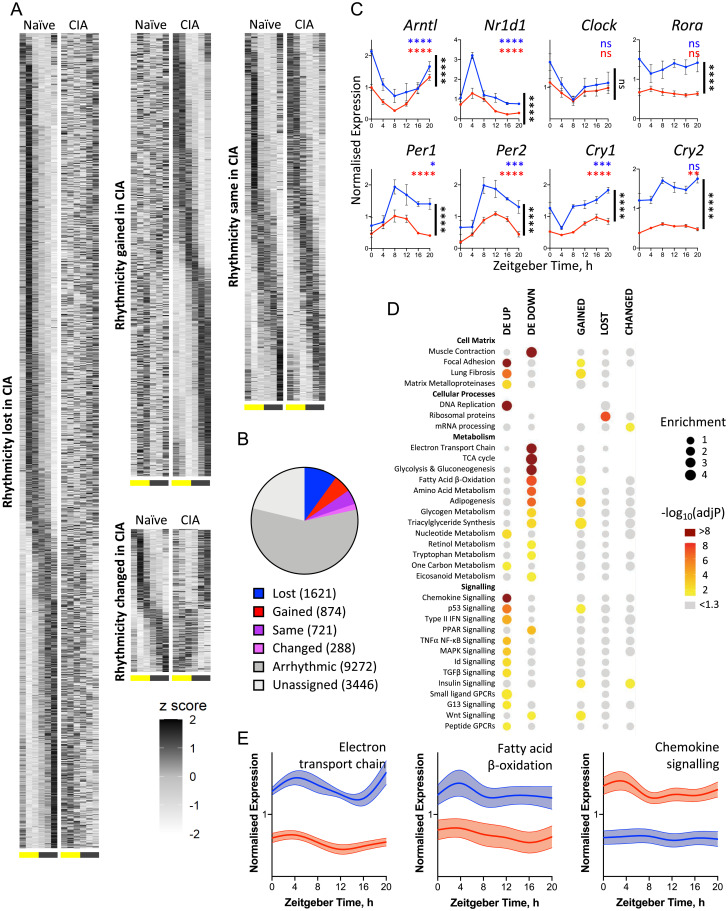
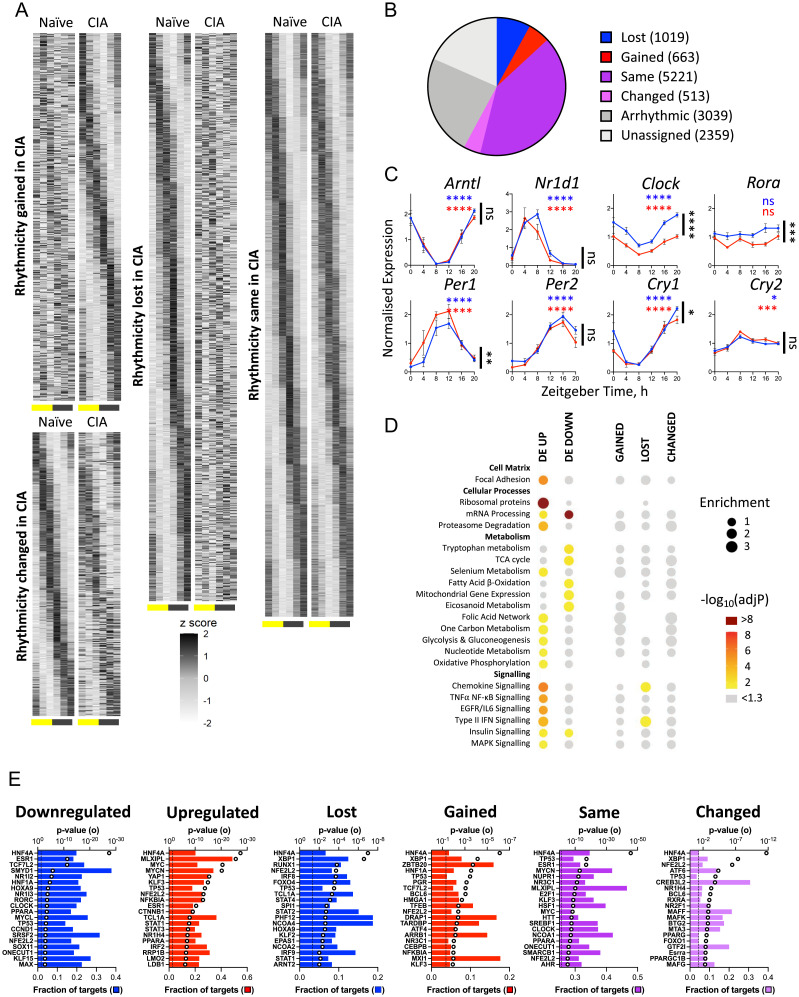
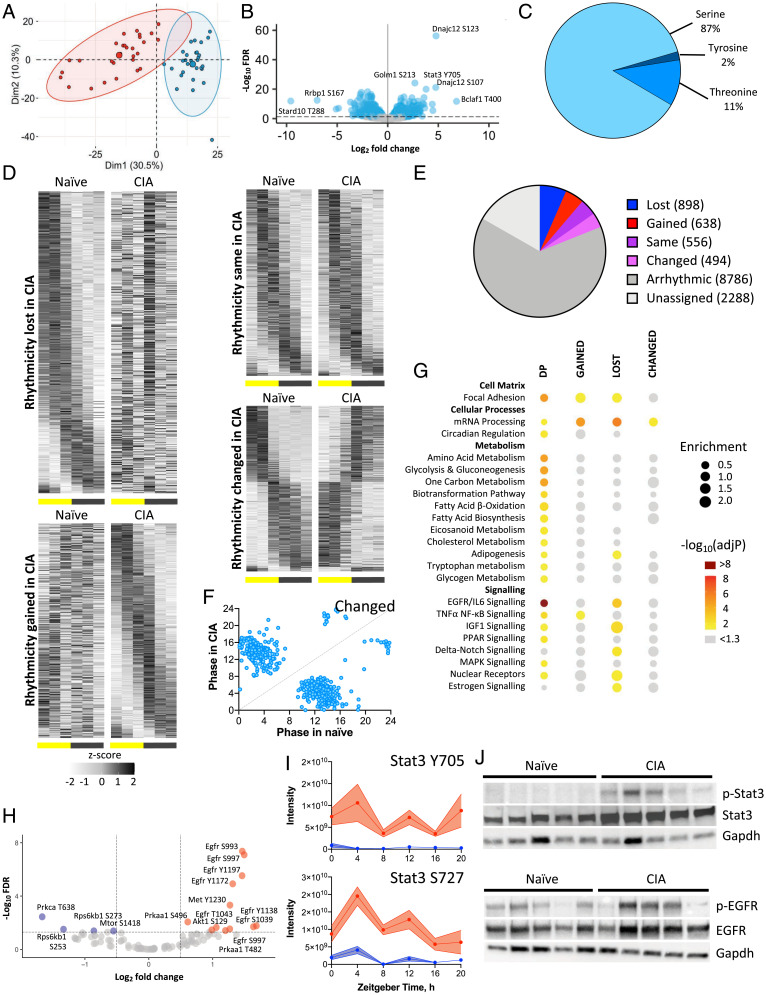
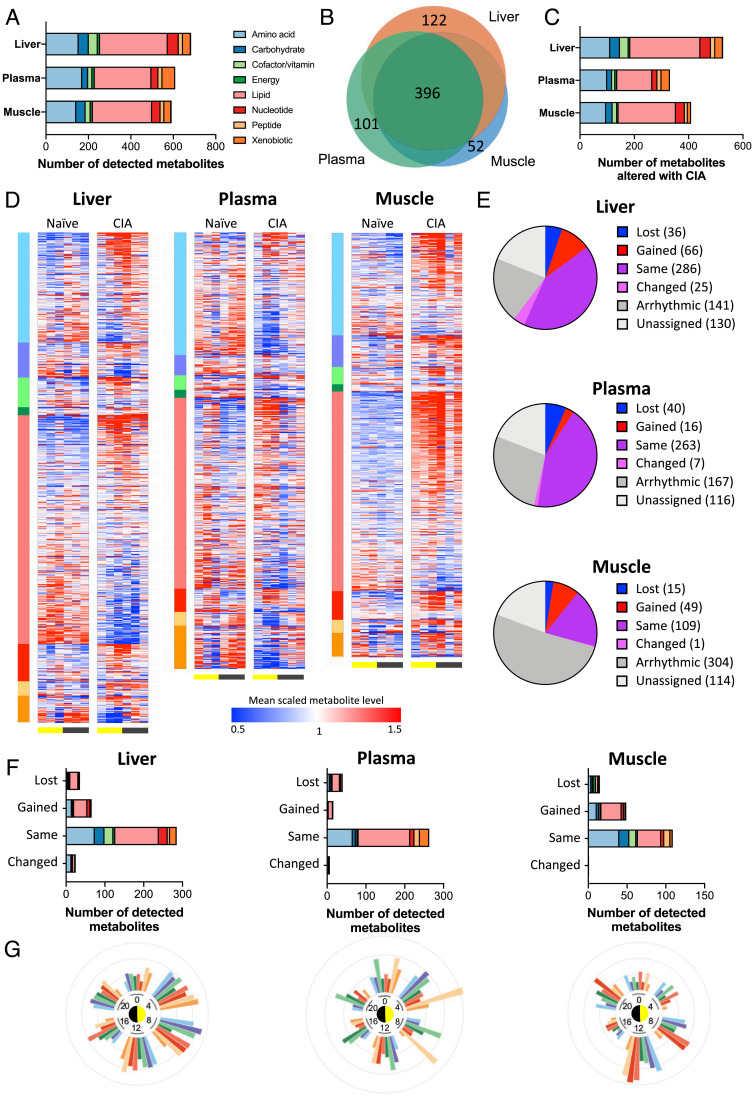
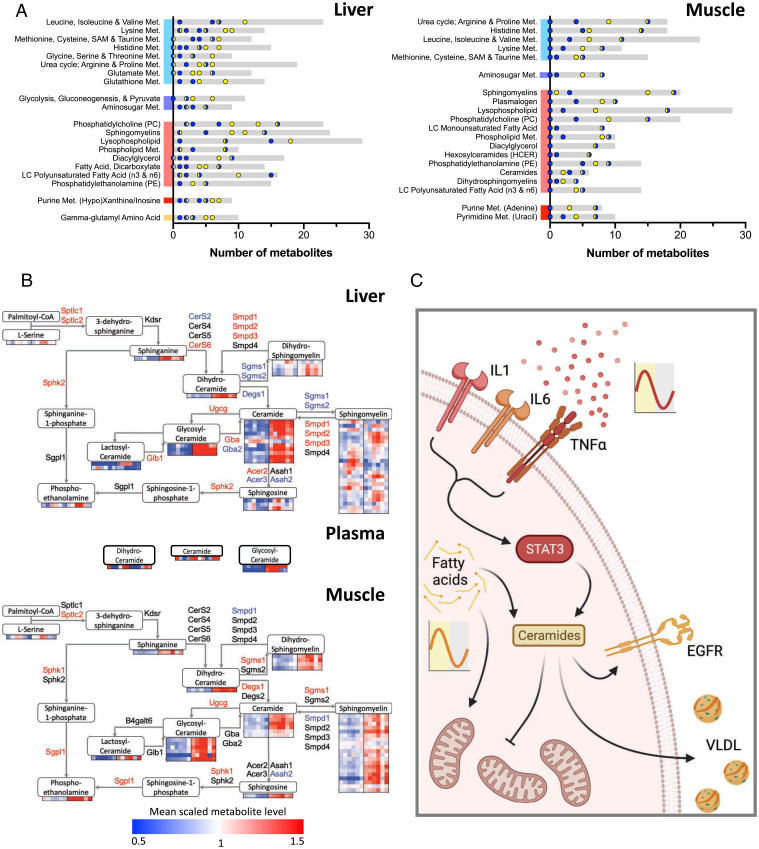
Chronic inflammation underpins many human diseases. Morbidity and mortality associated with chronic inflammation are often mediated through metabolic dysfunction. Inflammatory and metabolic processes vary through circadian time, suggesting an important temporal crosstalk between these systems. Using an established mouse model of rheumatoid arthritis, we show that chronic inflammatory arthritis results in rhythmic joint inflammation and drives major changes in muscle and liver energy metabolism and rhythmic gene expression. Transcriptional and phosphoproteomic analyses revealed alterations in lipid metabolism and mitochondrial function associated with increased EGFR-JAK-STAT3 signaling. Metabolomic analyses confirmed rhythmic metabolic rewiring with impaired β-oxidation and lipid handling and revealed a pronounced shunt toward sphingolipid and ceramide accumulation. The arthritis-related production of ceramides was most pronounced during the day, which is the time of peak inflammation and increased reliance on fatty acid oxidation. Thus, our data demonstrate that localized joint inflammation drives a time-of-day–dependent build-up of bioactive lipid species driven by rhythmic inflammation and altered EGFR-STAT signaling.
Keywords: ceramide; circadian clock; inflammation; mitochondria; rheumatoid arthritis.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Dibner C., Schibler U., Albrecht U., The mammalian circadian timing system: Organization and coordination of central and peripheral clocks. Annu. Rev. Physiol. 72, 517–549 (2010). - PubMed
-
- Bass J., Lazar M. A., Circadian time signatures of fitness and disease. Science 354, 994–999 (2016). - PubMed
-
- Scheiermann C., Gibbs J., Ince L., Loudon A., Clocking in to immunity. Nat. Rev. Immunol. 18, 423–437 (2018). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
