

Maladaptive innate immune training of myelopoiesis links inflammatory comorbidities
- PMID: 35483374
- PMCID: PMC9106933
- DOI: 10.1016/j.cell.2022.03.043
Maladaptive innate immune training of myelopoiesis links inflammatory comorbidities
Abstract
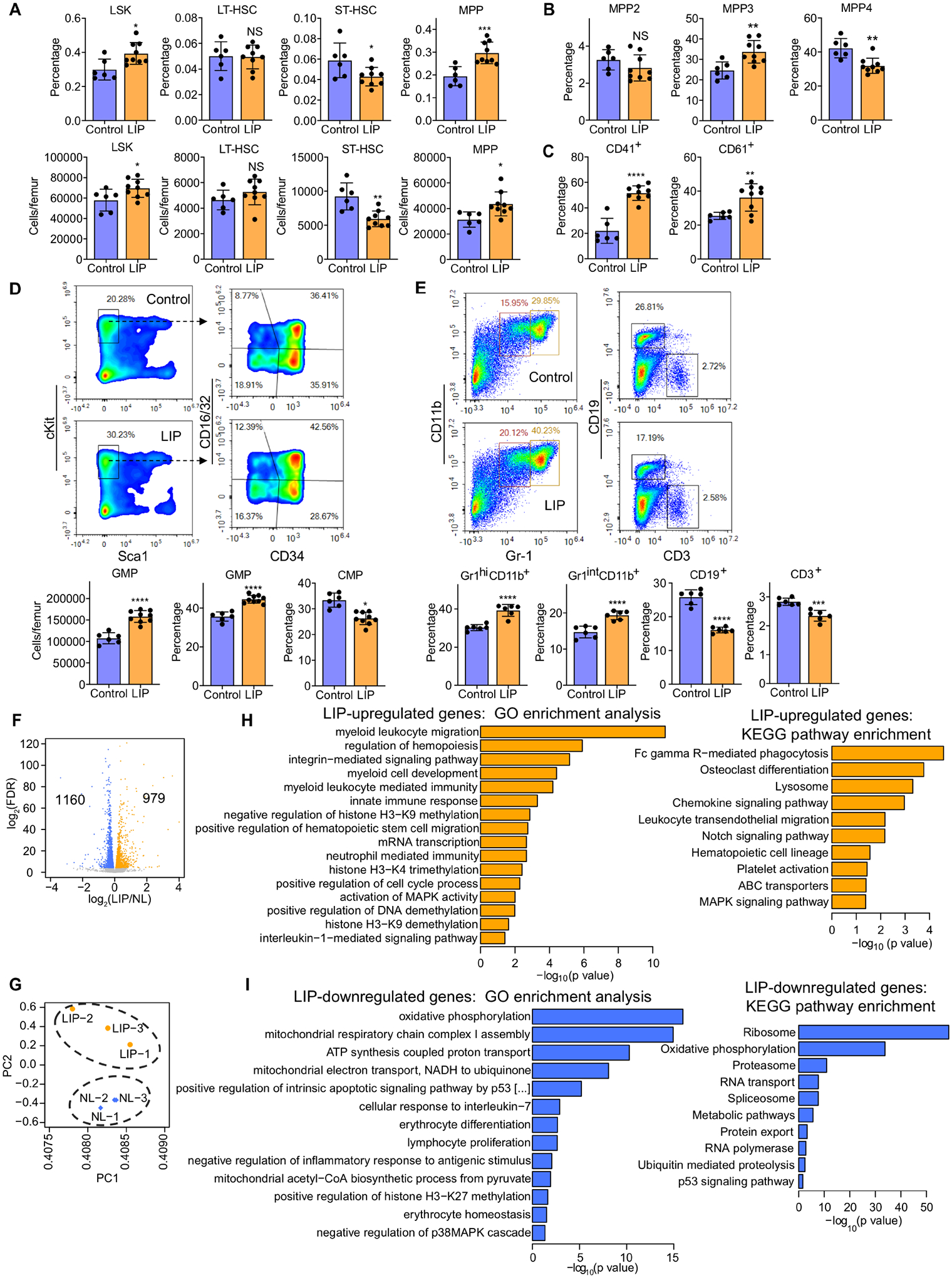
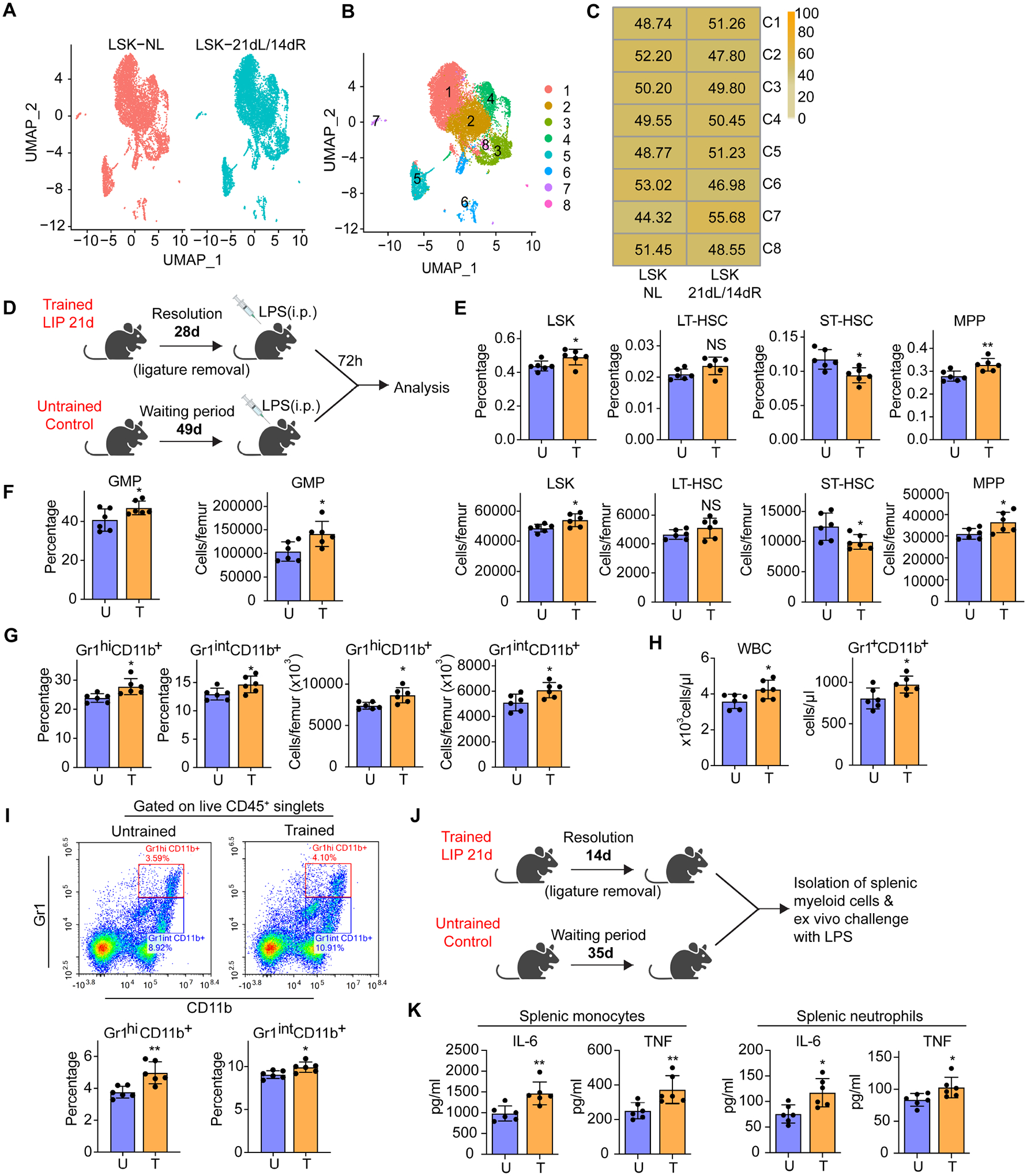
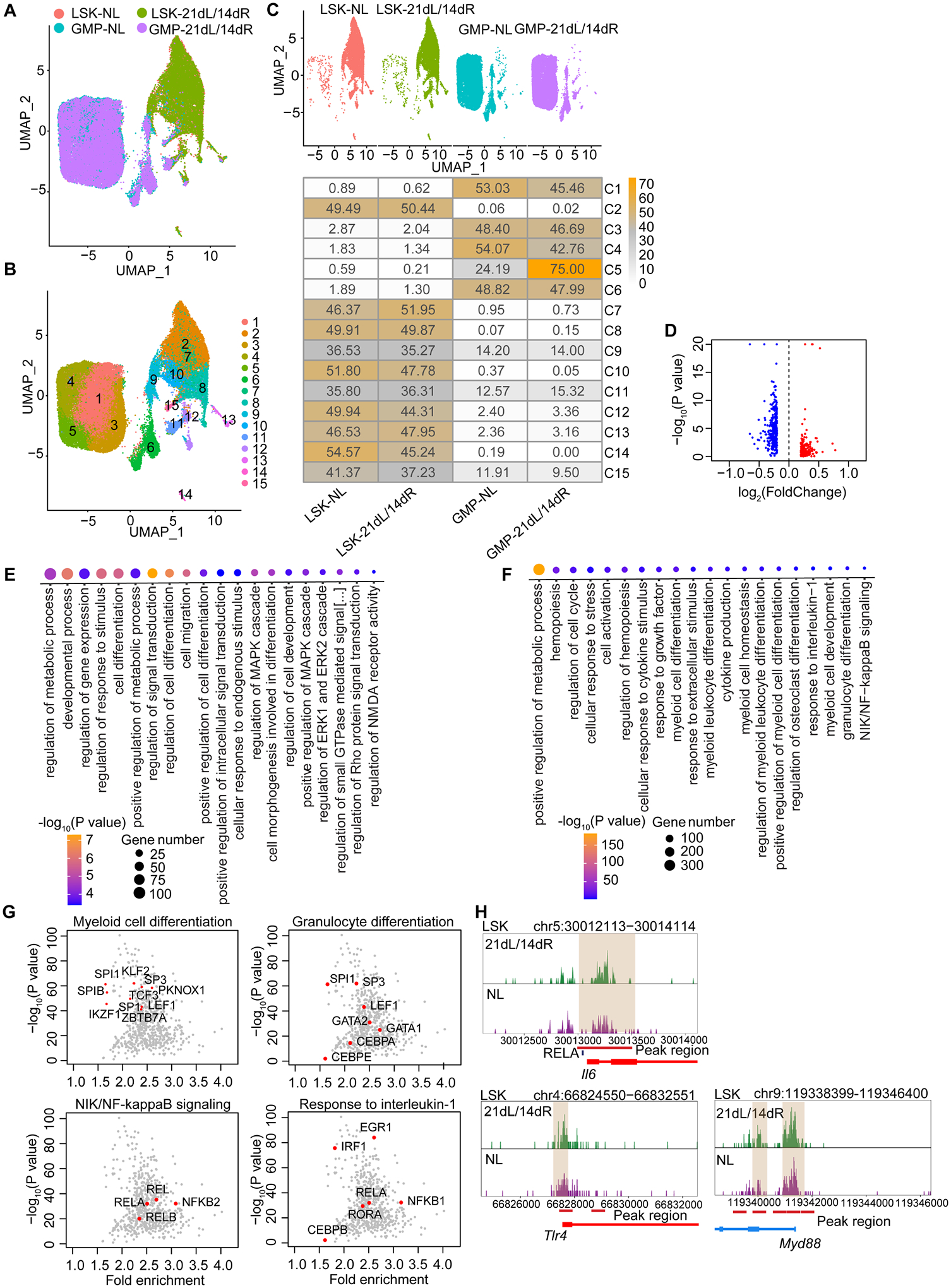
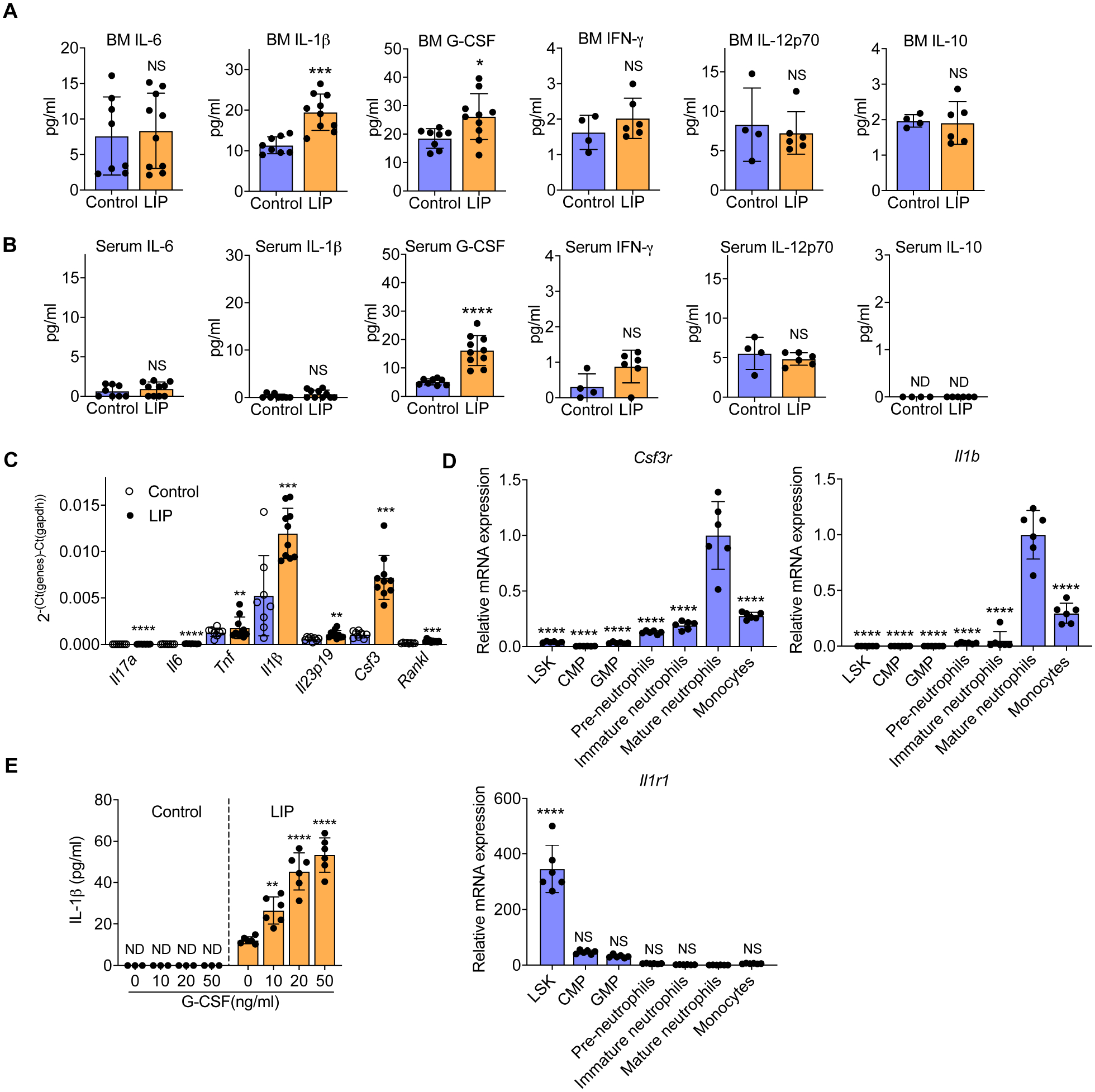
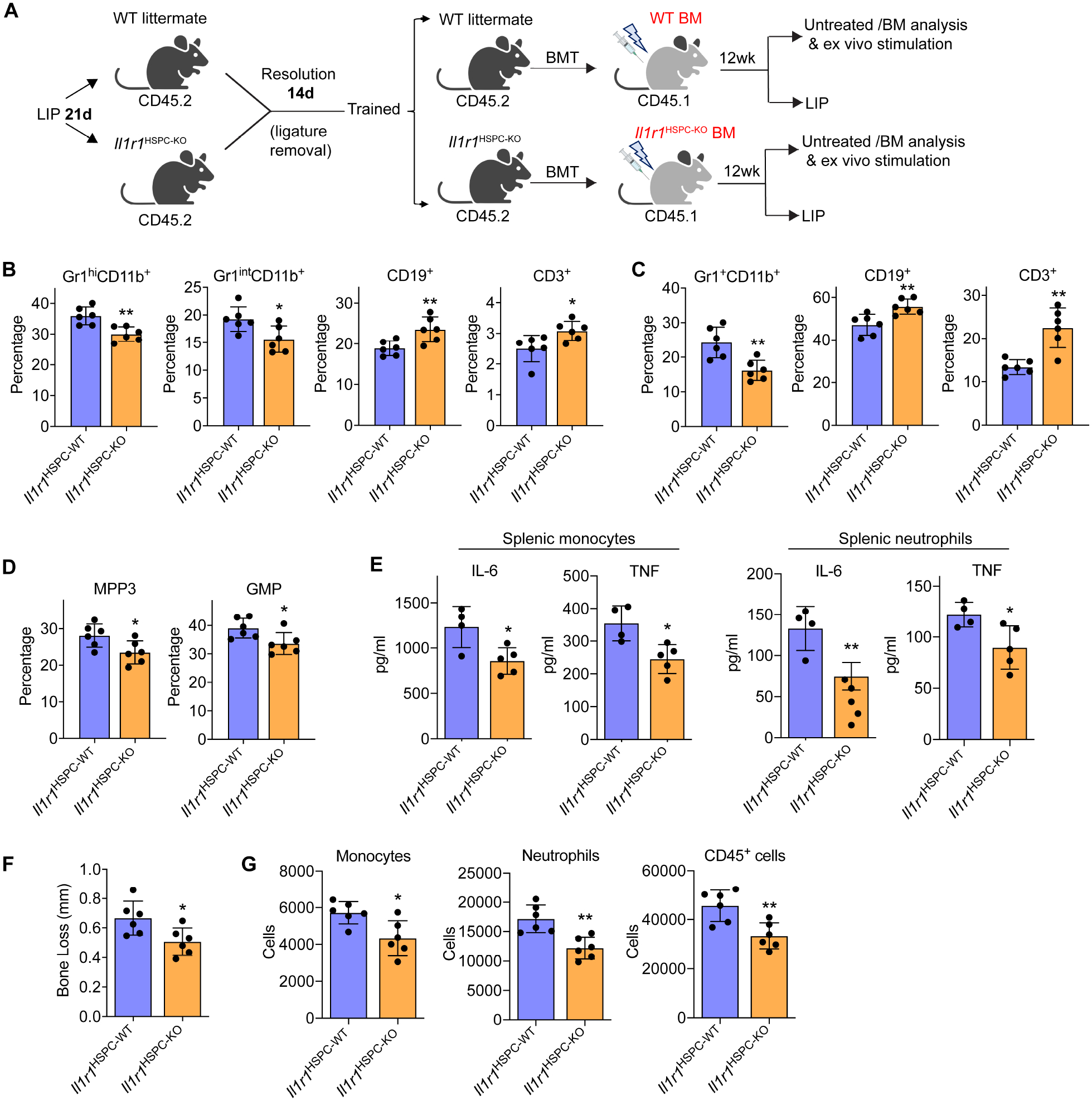
Bone marrow (BM)-mediated trained innate immunity (TII) is a state of heightened immune responsiveness of hematopoietic stem and progenitor cells (HSPC) and their myeloid progeny. We show here that maladaptive BM-mediated TII underlies inflammatory comorbidities, as exemplified by the periodontitis-arthritis axis. Experimental-periodontitis-related systemic inflammation in mice induced epigenetic rewiring of HSPC and led to sustained enhancement of production of myeloid cells with increased inflammatory preparedness. The periodontitis-induced trained phenotype was transmissible by BM transplantation to naive recipients, which exhibited increased inflammatory responsiveness and disease severity when subjected to inflammatory arthritis. IL-1 signaling in HSPC was essential for their maladaptive training by periodontitis. Therefore, maladaptive innate immune training of myelopoiesis underlies inflammatory comorbidities and may be pharmacologically targeted to treat them via a holistic approach.
Keywords: arthritis; bone marrow transplantation; comorbidities; epigenetic rewiring; hematopoietic stem and progenitor cells; inflammation; myelopoiesis; periodontitis; trained immunity.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests M.G.N. is the scientific founder and member of the Scientific Advisory Board of Trained Therapeutics and Discovery (TTxD). M.G.N. has two patents: US18/61935 (Targeted nanoimmunotherapy to increase trained immunity) and US18/61939 (Targeted nanoimmunotherapy for inhibition of trained immunity). The other authors declare no competing interests.
Figures


Comment in
-
Fired up in the bone marrow.Nat Rev Immunol. 2022 Jun;22(6):336-337. doi: 10.1038/s41577-022-00733-0. Nat Rev Immunol. 2022. PMID: 35508808 No abstract available.
-
Inflammatory comorbidities: to train or not to train?Trends Immunol. 2022 Jun;43(6):420-422. doi: 10.1016/j.it.2022.04.005. Epub 2022 May 8. Trends Immunol. 2022. PMID: 35537982
References
-
- Berendsen MLT, Schaltz-Buchholzer F, Bles P, Biering-Sørensen S, Jensen KJ, Monteiro I, Silva I, Aaby P, and Benn CS (2021). Parental Bacillus Calmette-Guérin vaccine scars decrease infant mortality in the first six weeks of life: A retrospective cohort study. EClinicalMedicine 39, 101049. - PMC - PubMed
-
- Bernelot Moens SJ, van der Valk FM, Strang AC, Kroon J, Smits LP, Kneepkens EL, Verberne HJ, van Buul JD, Nurmohamed MT, and Stroes ES (2016). Unexpected arterial wall and cellular inflammation in patients with rheumatoid arthritis in remission using biological therapy: a cross-sectional study. Arthritis Res Ther 18, 115. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
