

Gut microbial β-glucuronidases regulate host luminal proteases and are depleted in irritable bowel syndrome
- PMID: 35484230
- PMCID: PMC9081267
- DOI: 10.1038/s41564-022-01103-1
Gut microbial β-glucuronidases regulate host luminal proteases and are depleted in irritable bowel syndrome
Abstract
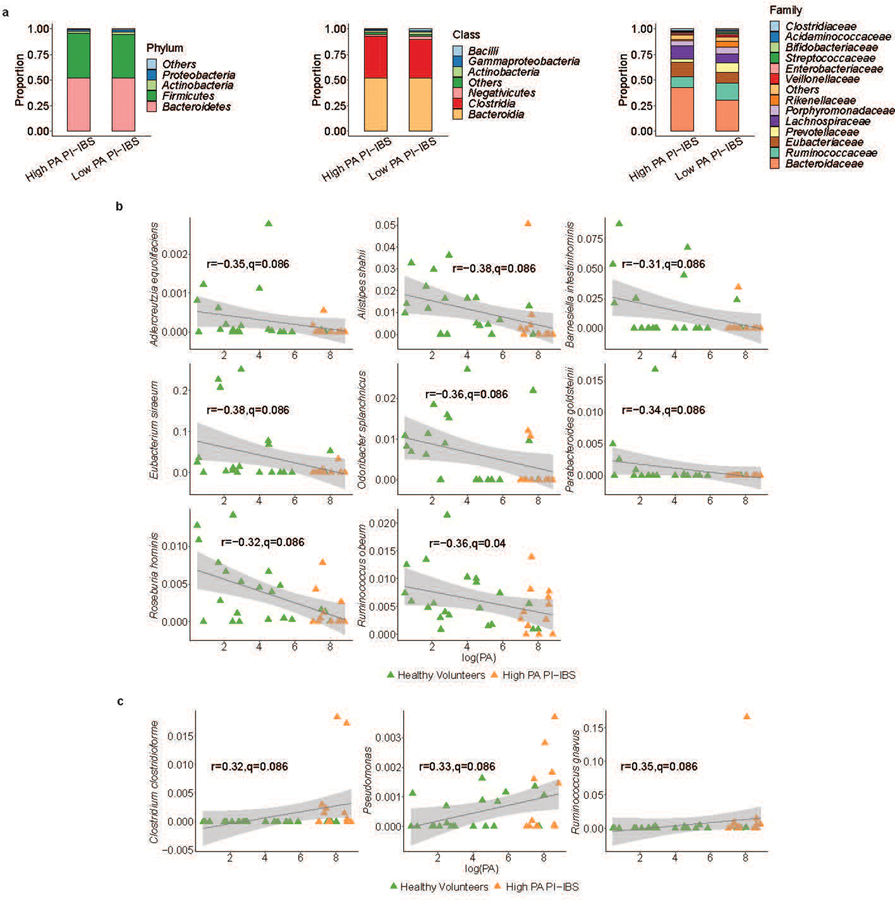
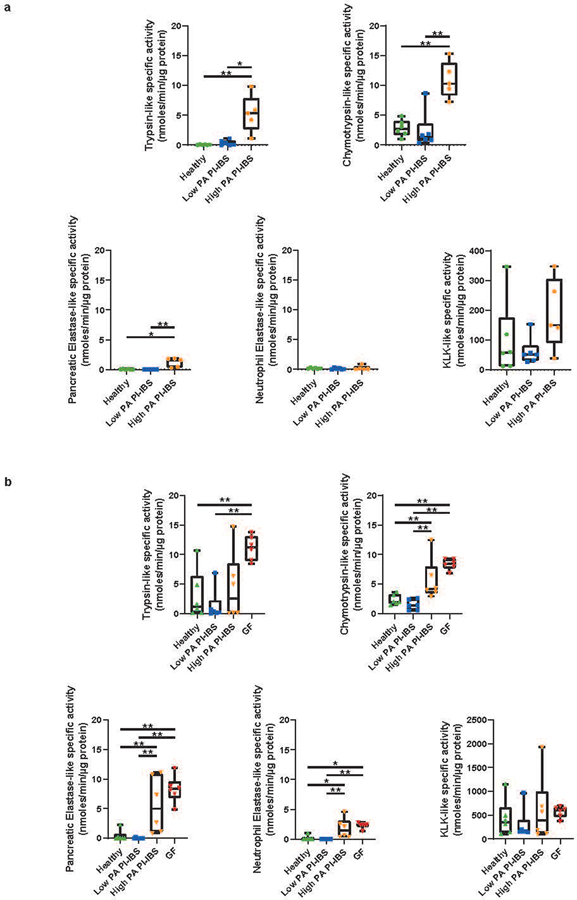
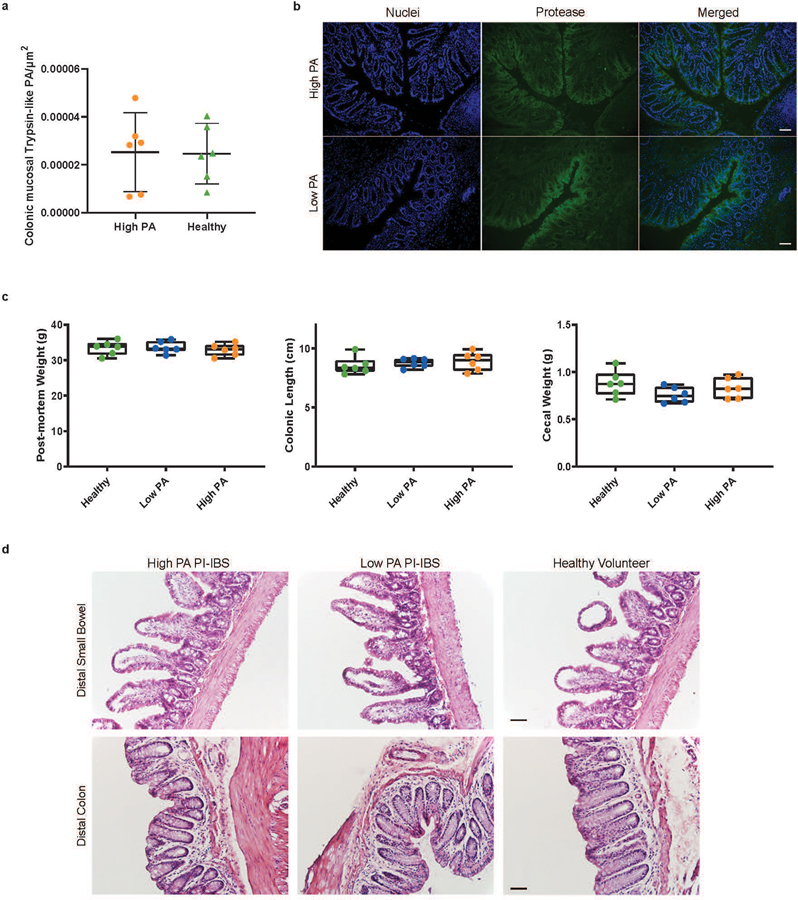
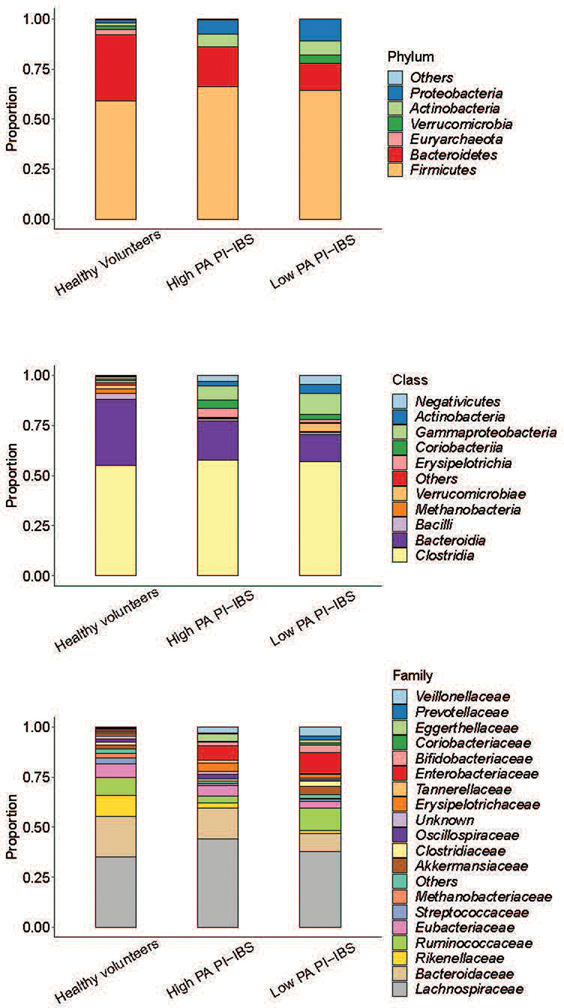
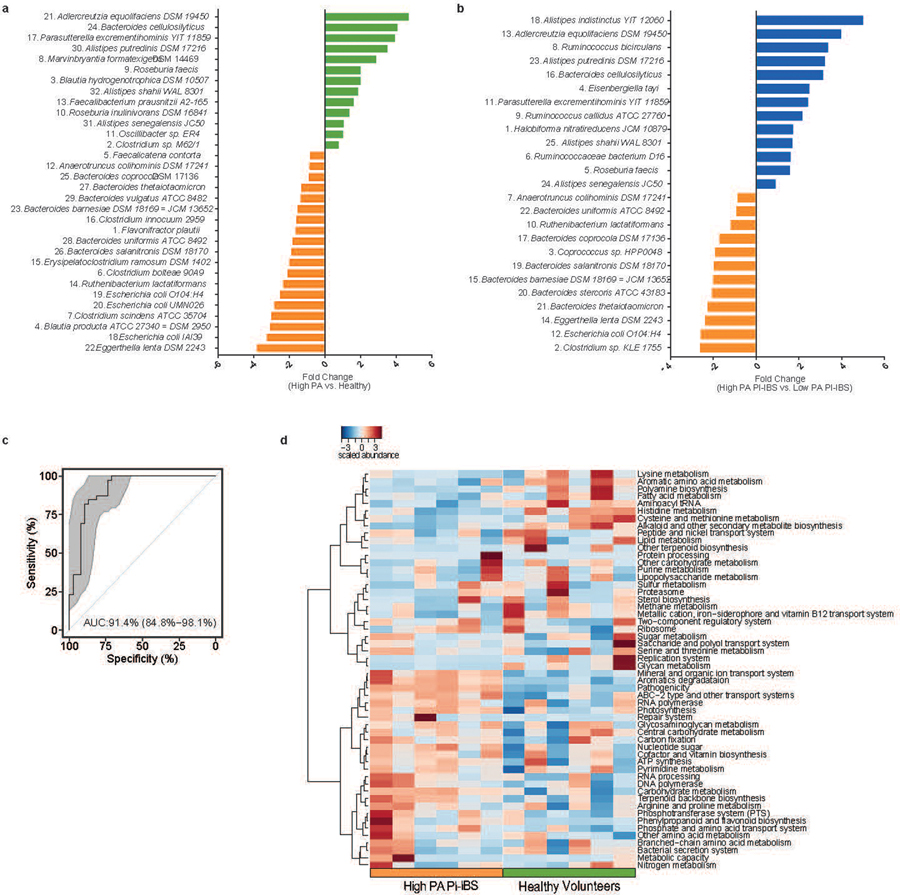
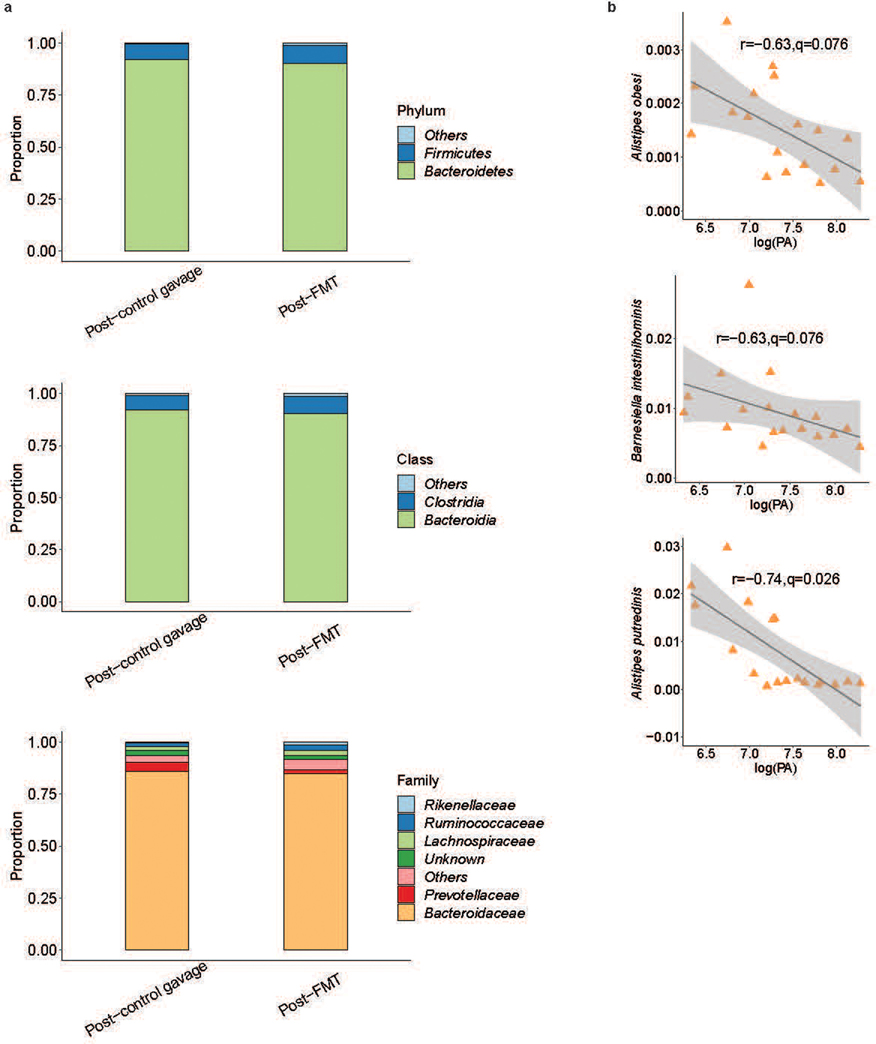
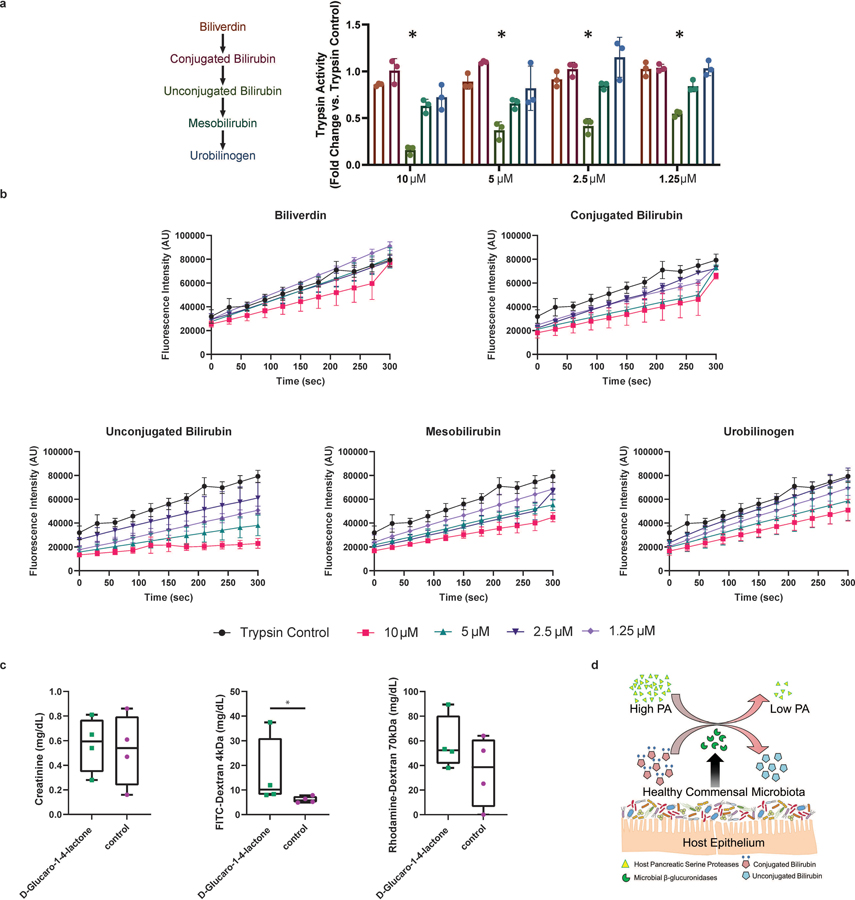
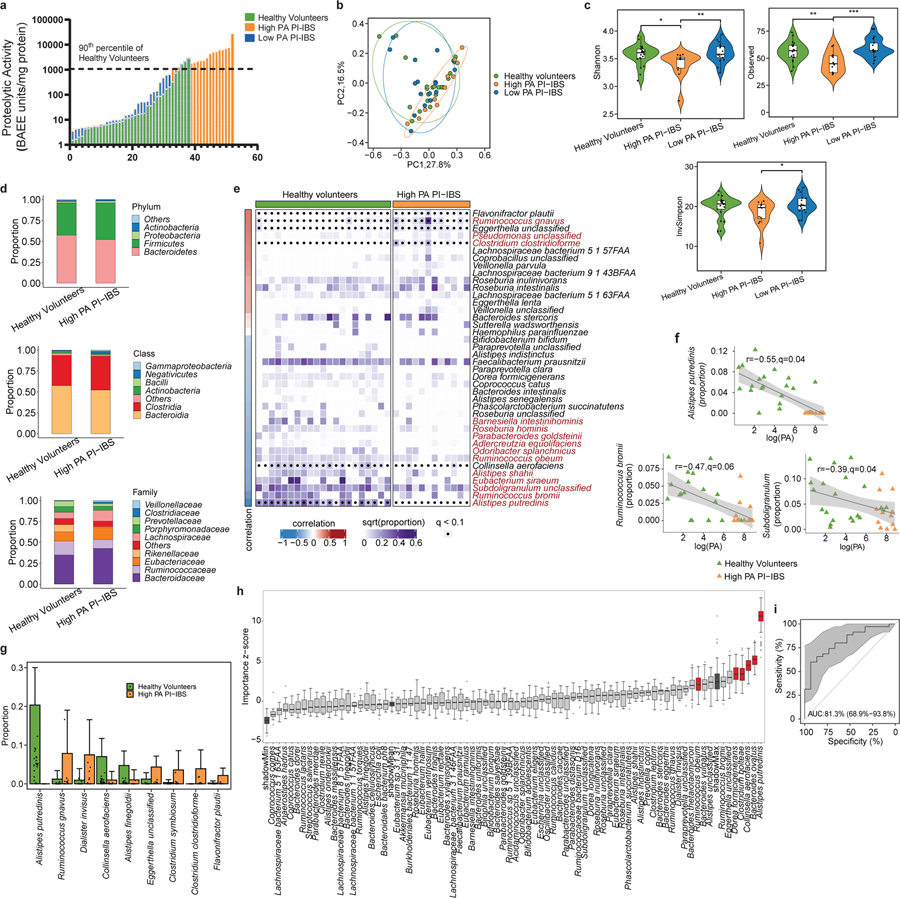
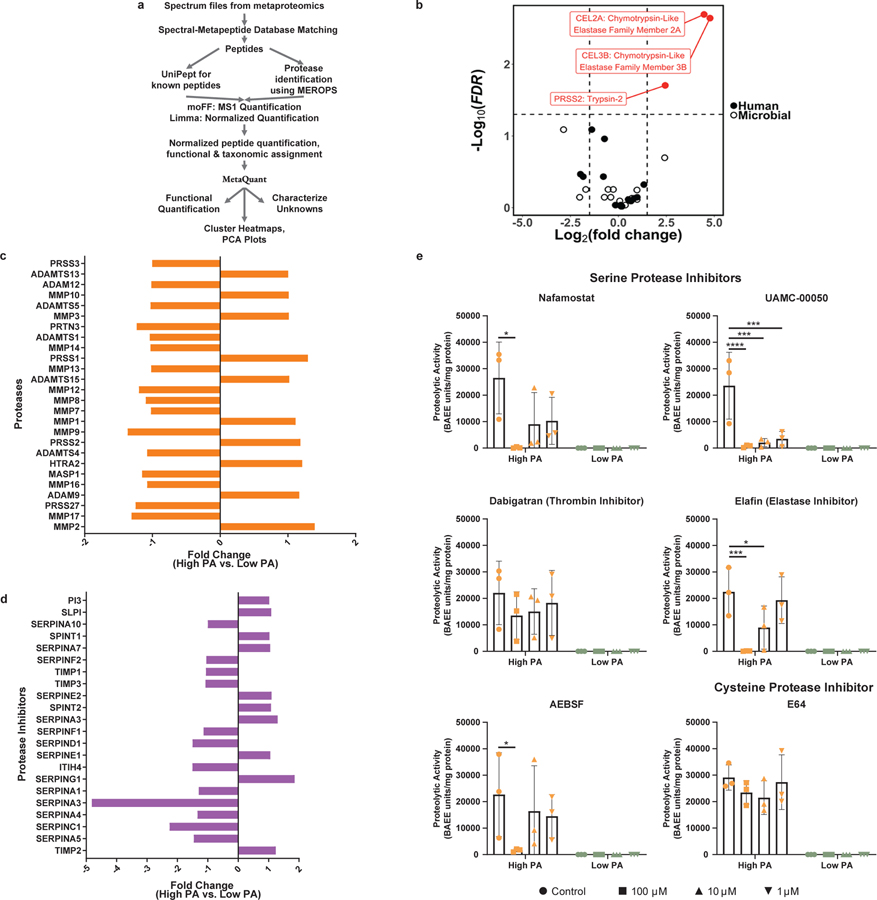
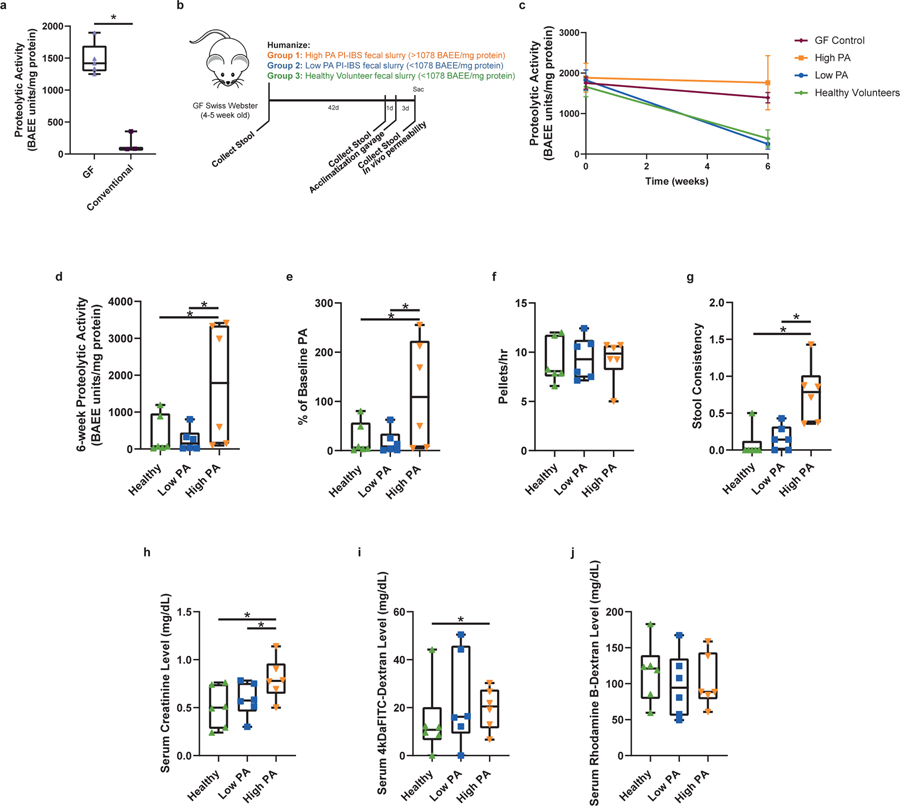
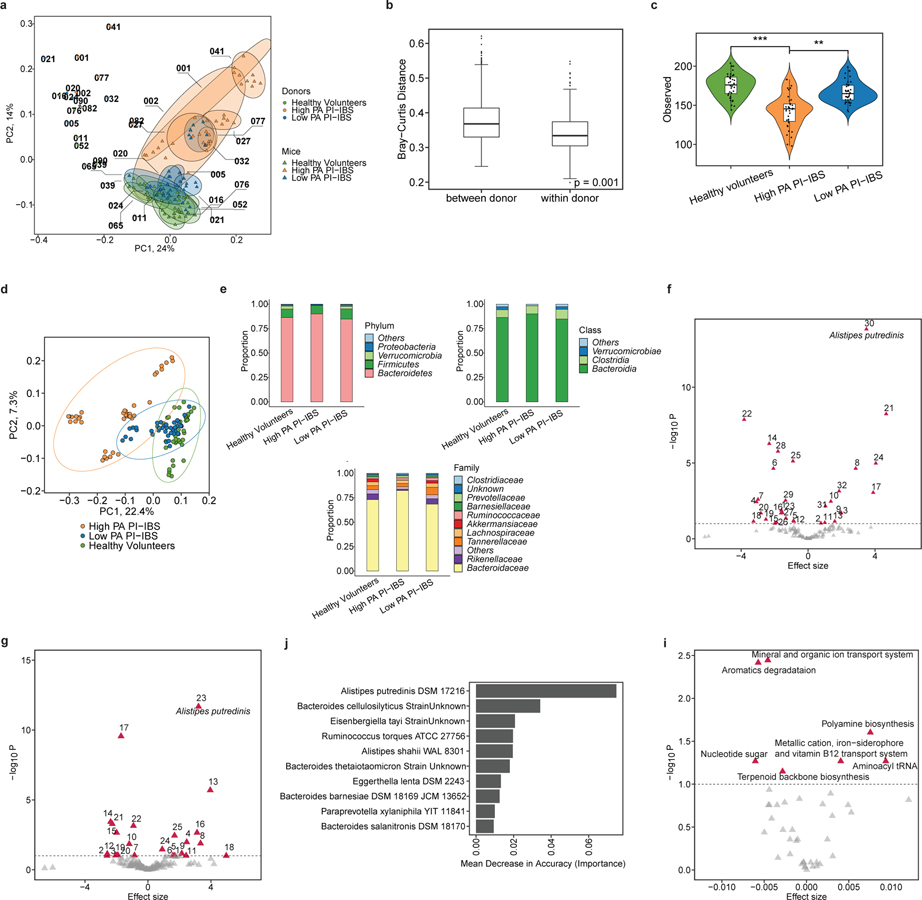
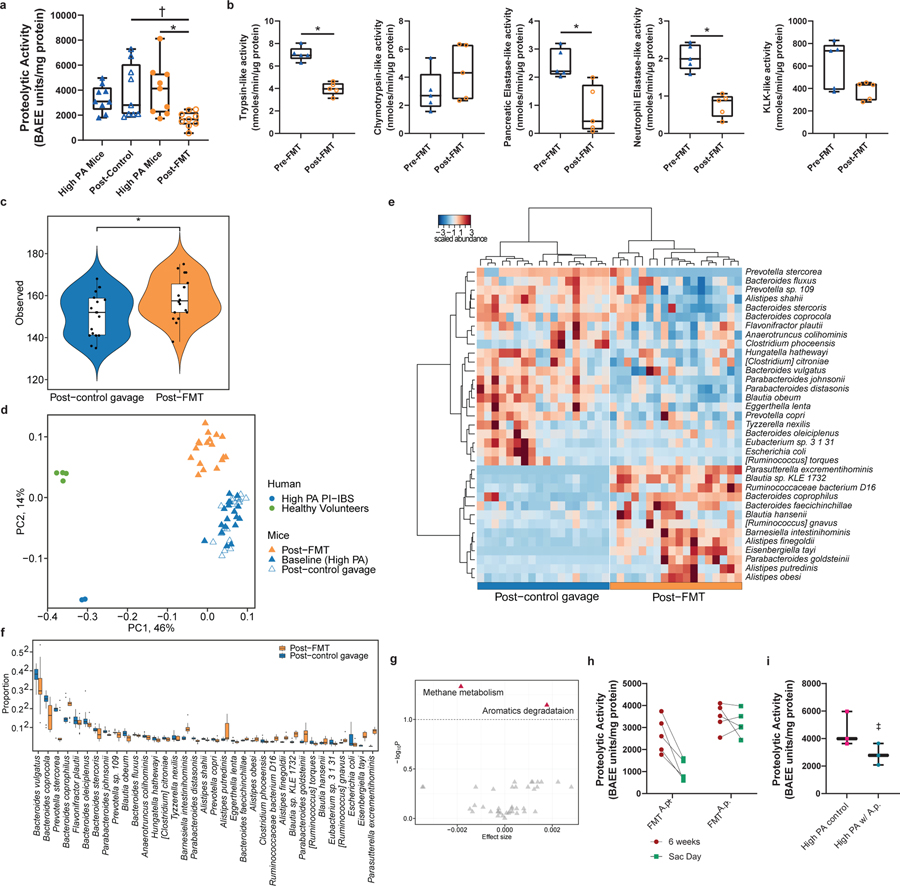
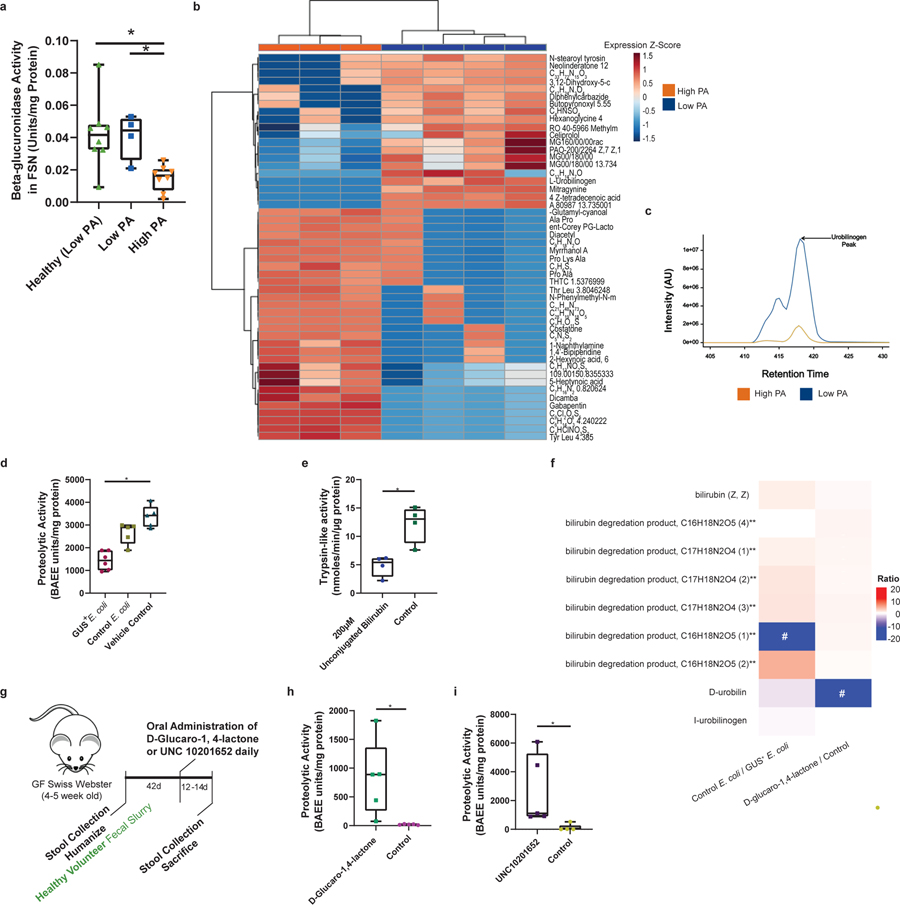
Intestinal proteases mediate digestion and immune signalling, while increased gut proteolytic activity disrupts the intestinal barrier and generates visceral hypersensitivity, which is common in irritable bowel syndrome (IBS). However, the mechanisms controlling protease function are unclear. Here we show that members of the gut microbiota suppress intestinal proteolytic activity through production of unconjugated bilirubin. This occurs via microbial β-glucuronidase-mediated conversion of bilirubin conjugates. Metagenomic analysis of faecal samples from patients with post-infection IBS (n = 52) revealed an altered gut microbiota composition, in particular a reduction in Alistipes taxa, and high gut proteolytic activity driven by specific host serine proteases compared with controls. Germ-free mice showed 10-fold higher proteolytic activity compared with conventional mice. Colonization with microbiota samples from high proteolytic activity IBS patients failed to suppress proteolytic activity in germ-free mice, but suppression of proteolytic activity was achieved with colonization using microbiota from healthy donors. High proteolytic activity mice had higher intestinal permeability, a higher relative abundance of Bacteroides and a reduction in Alistipes taxa compared with low proteolytic activity mice. High proteolytic activity IBS patients had lower fecal β-glucuronidase activity and end-products of bilirubin deconjugation. Mice treated with unconjugated bilirubin and β-glucuronidase-overexpressing E. coli significantly reduced proteolytic activity, while inhibitors of microbial β-glucuronidases increased proteolytic activity. Together, these data define a disease-relevant mechanism of host-microbial interaction that maintains protease homoeostasis in the gut.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Figures
Comment in
-
Regulating protease homeostasis in irritable bowel syndrome.Nat Rev Gastroenterol Hepatol. 2022 Jul;19(7):415. doi: 10.1038/s41575-022-00629-3. Nat Rev Gastroenterol Hepatol. 2022. PMID: 35524133 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
