

Screening the Toxoplasma kinome with high-throughput tagging identifies a regulator of invasion and egress
- PMID: 35484233
- PMCID: PMC9167752
- DOI: 10.1038/s41564-022-01104-0
Screening the Toxoplasma kinome with high-throughput tagging identifies a regulator of invasion and egress
Abstract
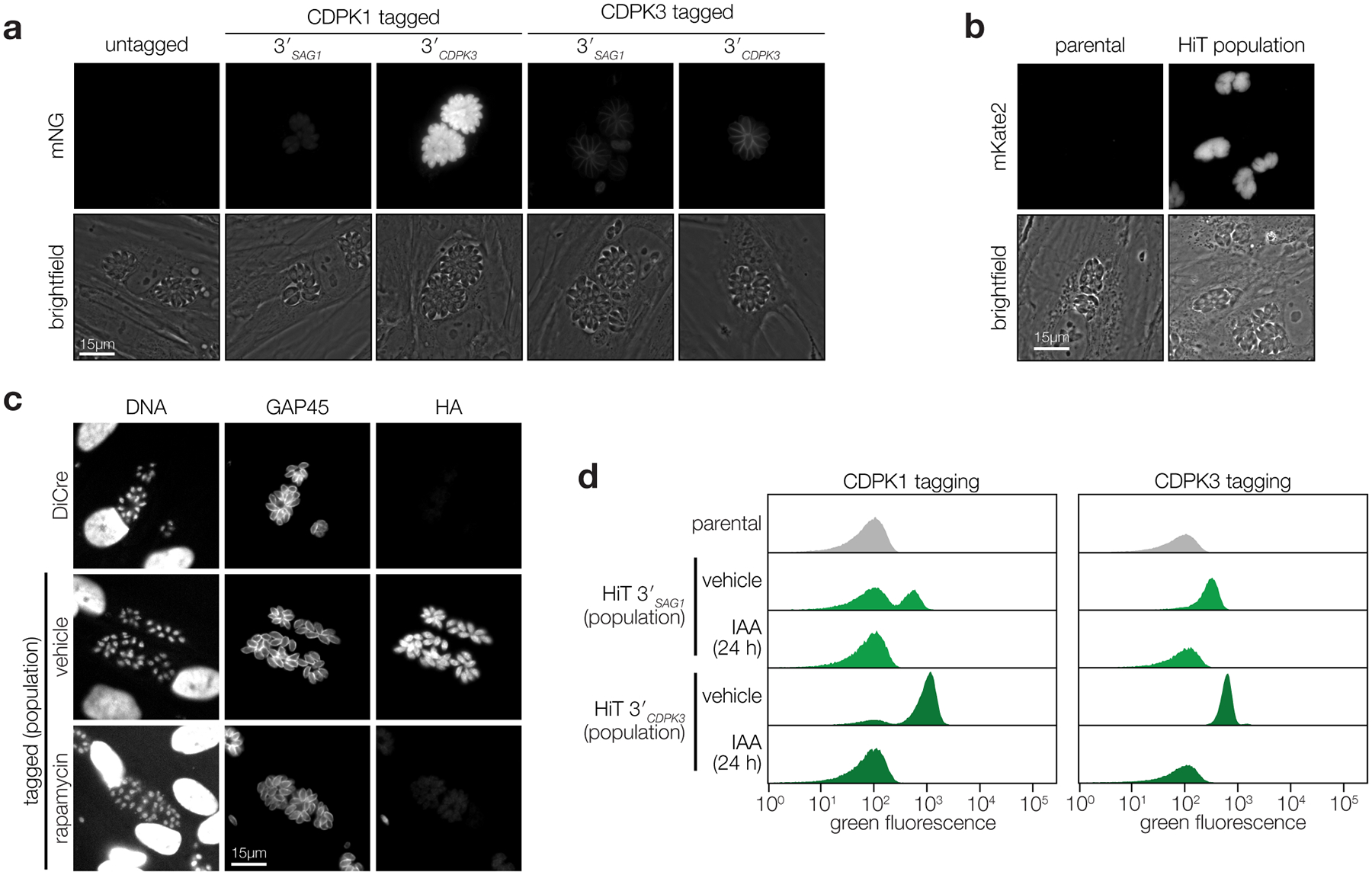
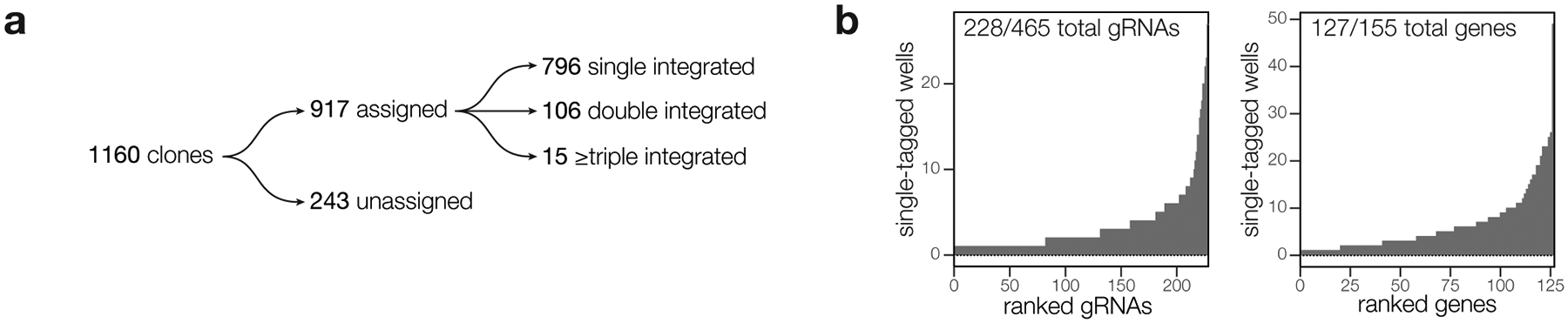
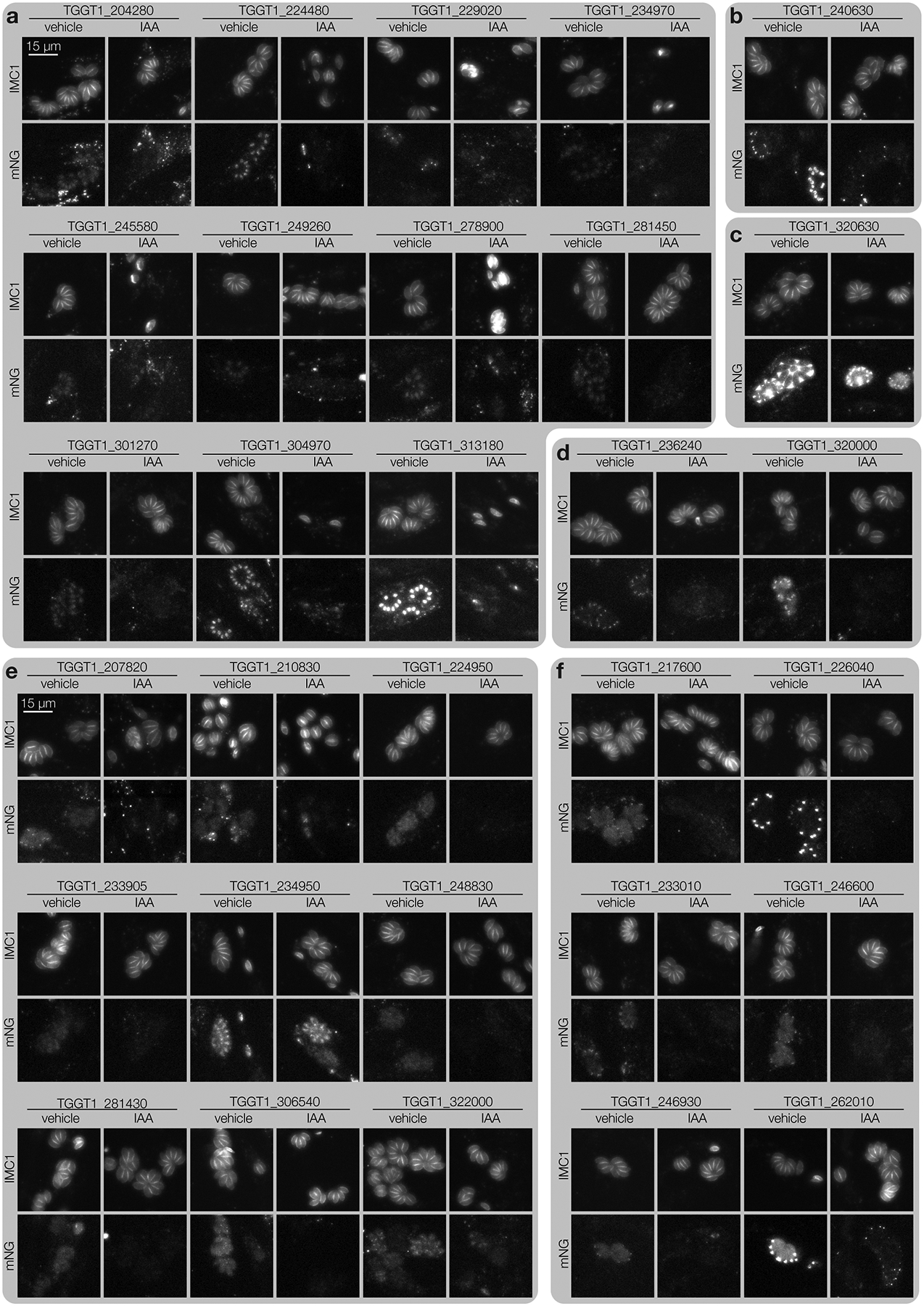
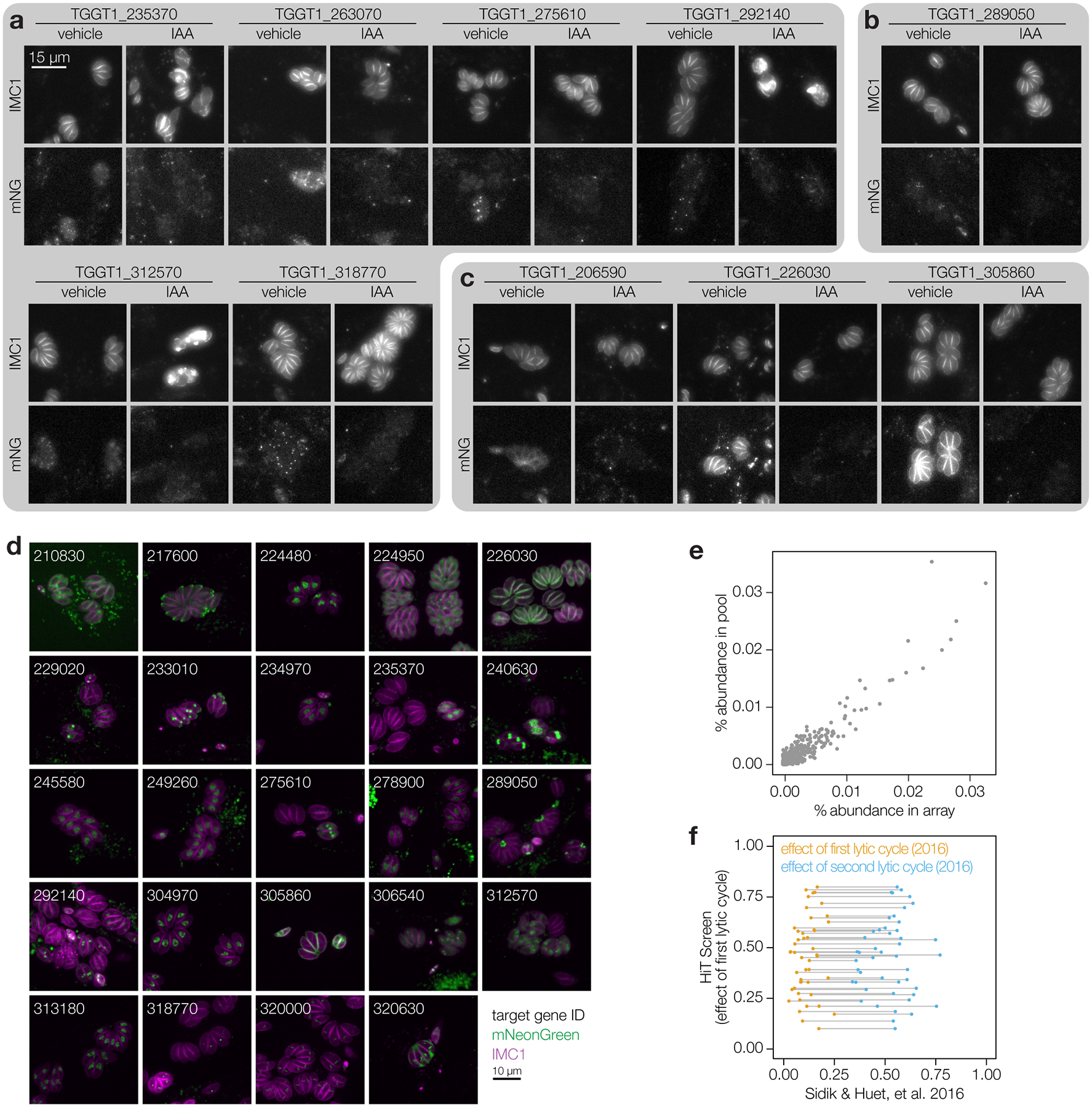
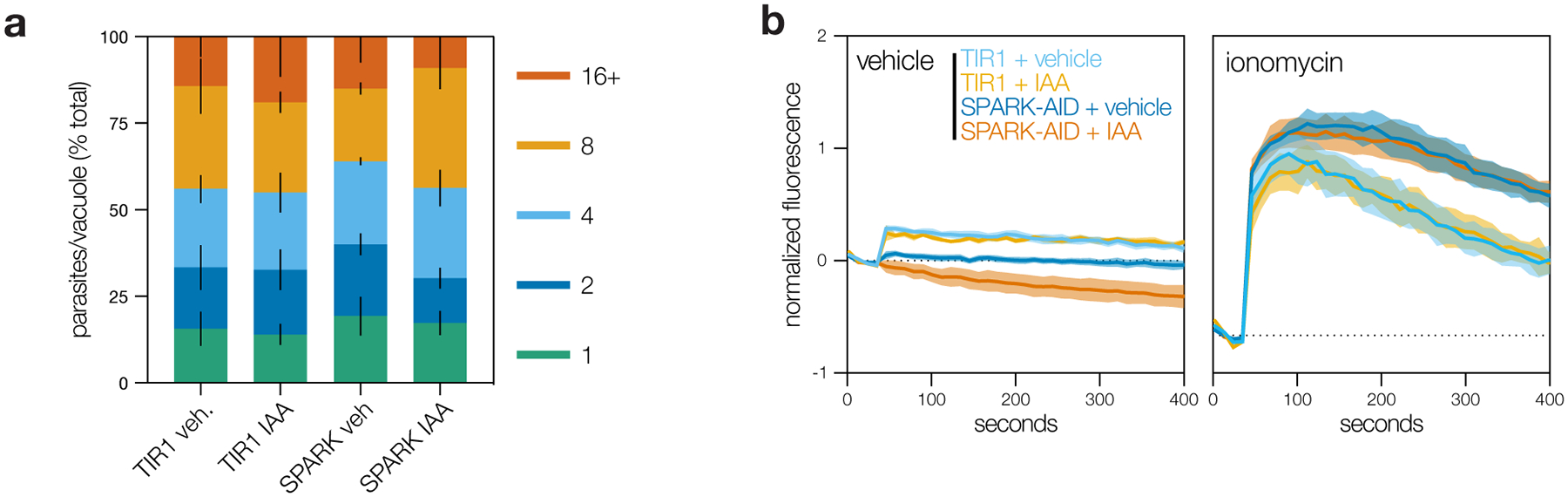
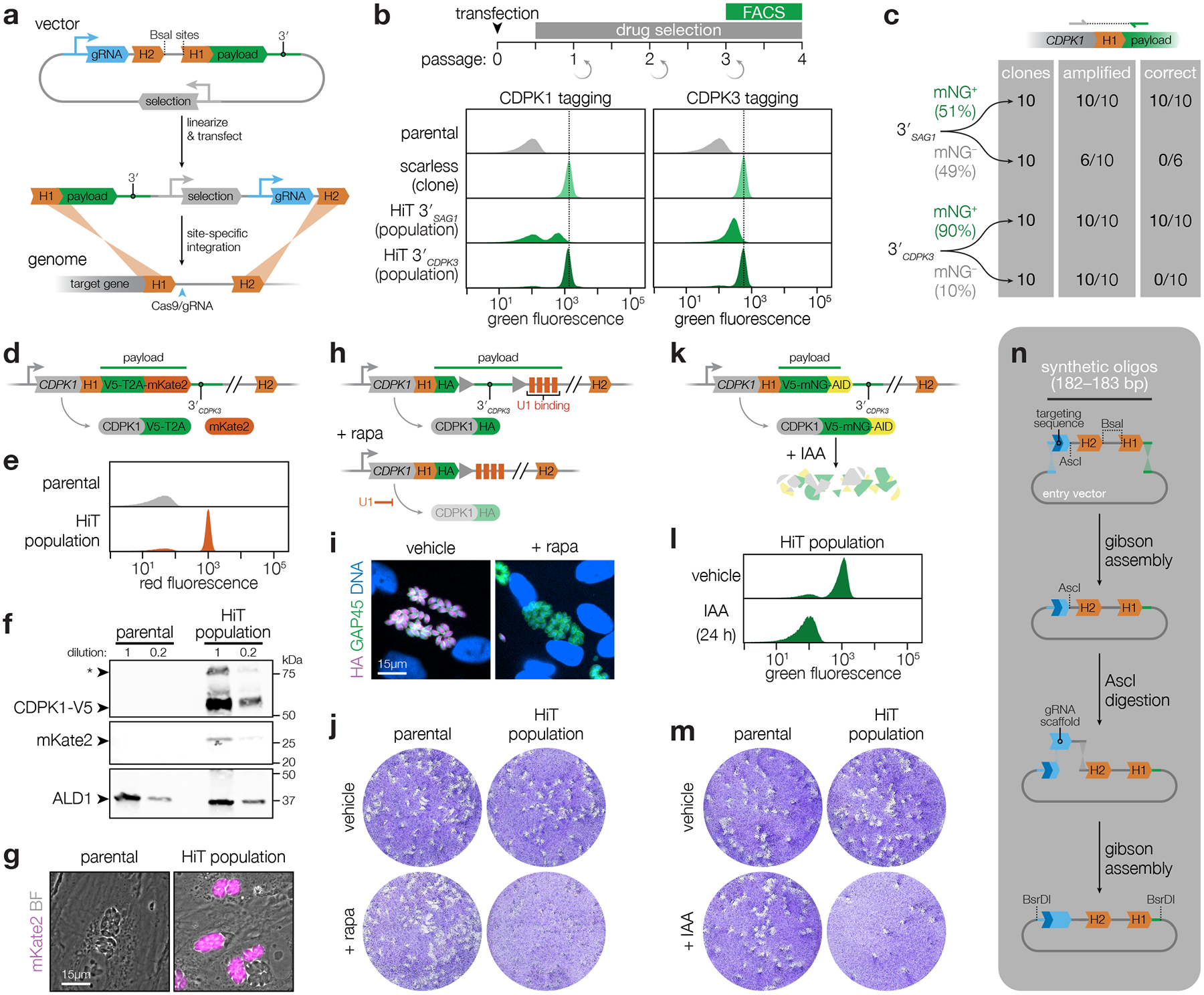
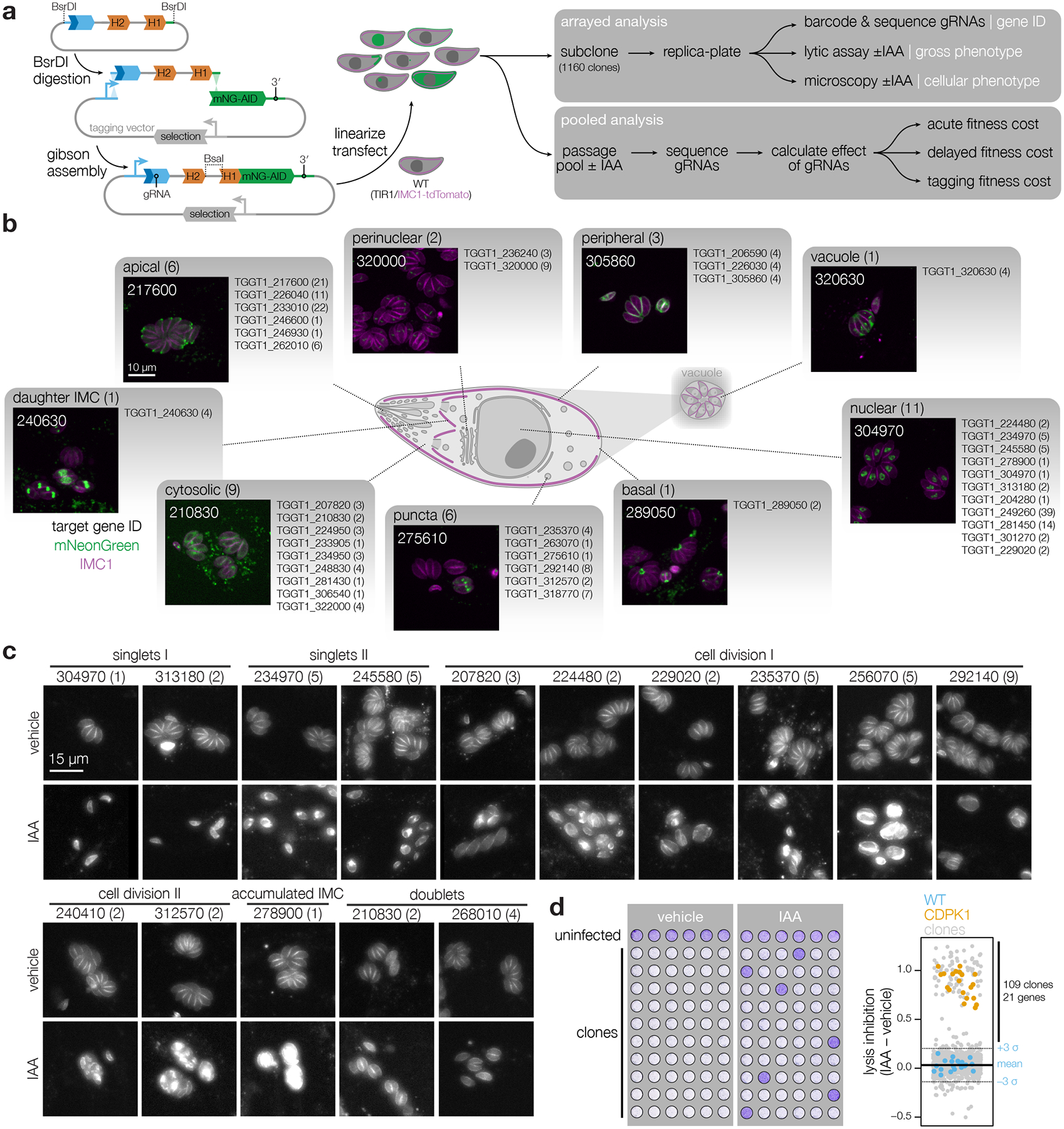
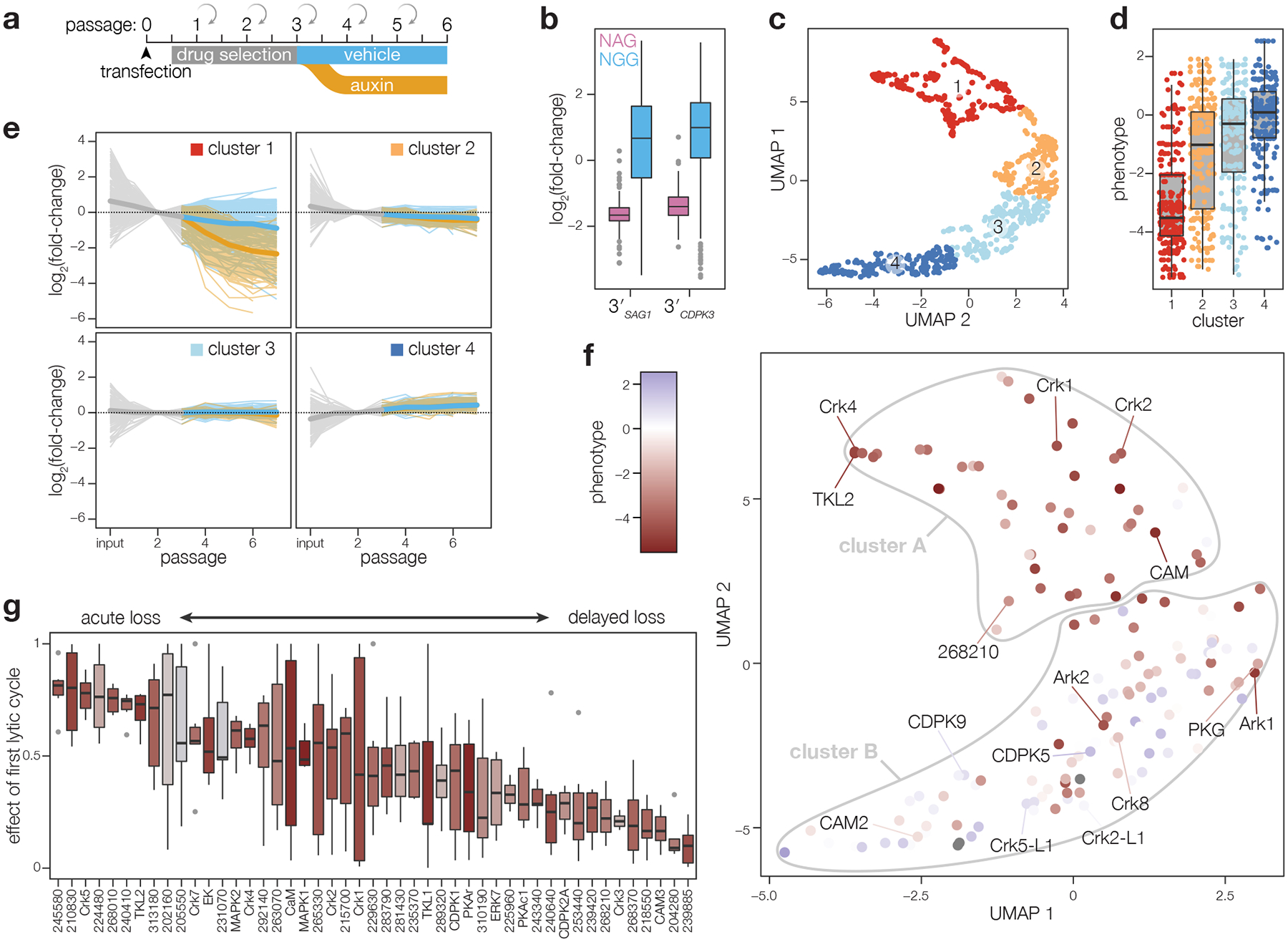
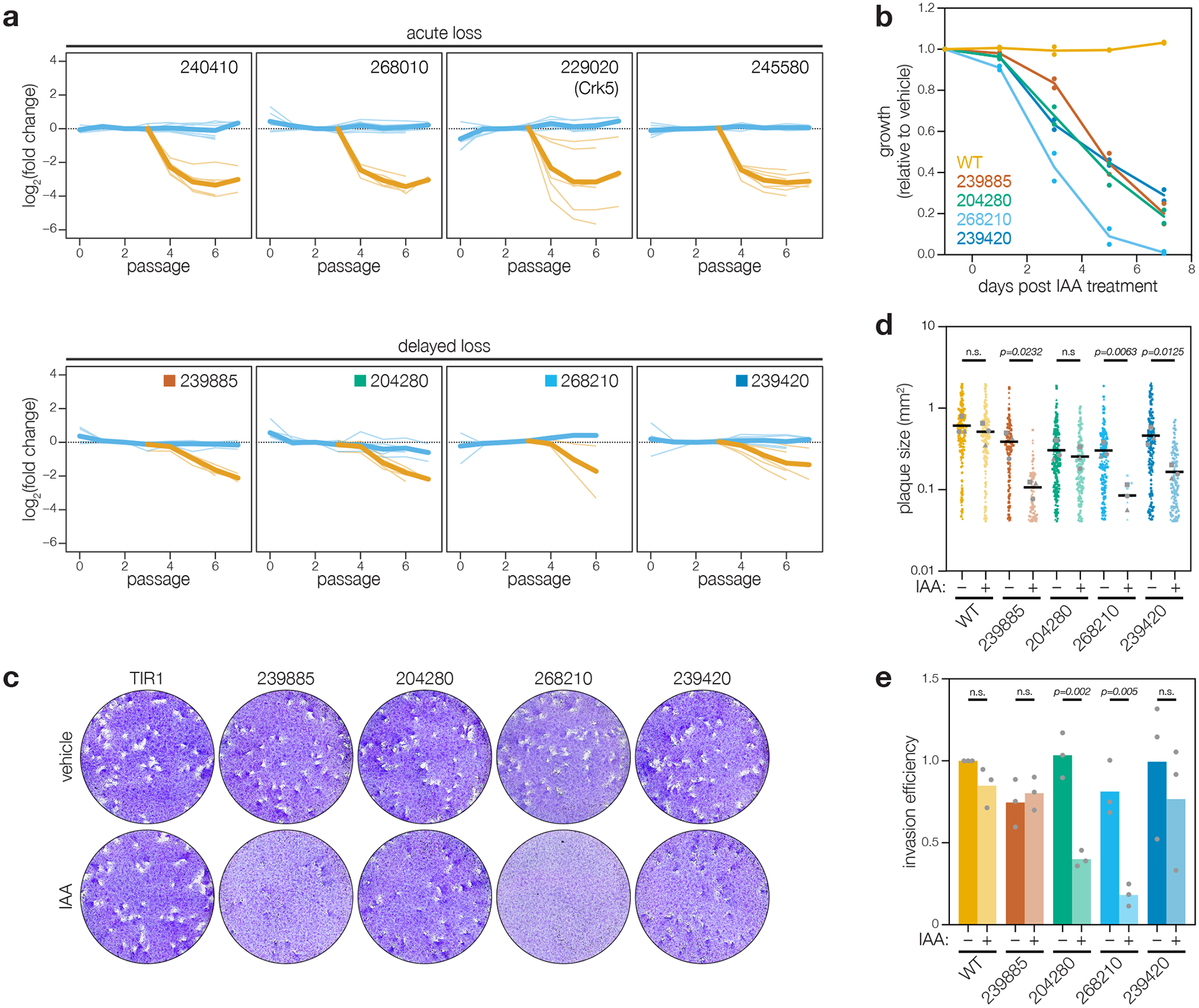
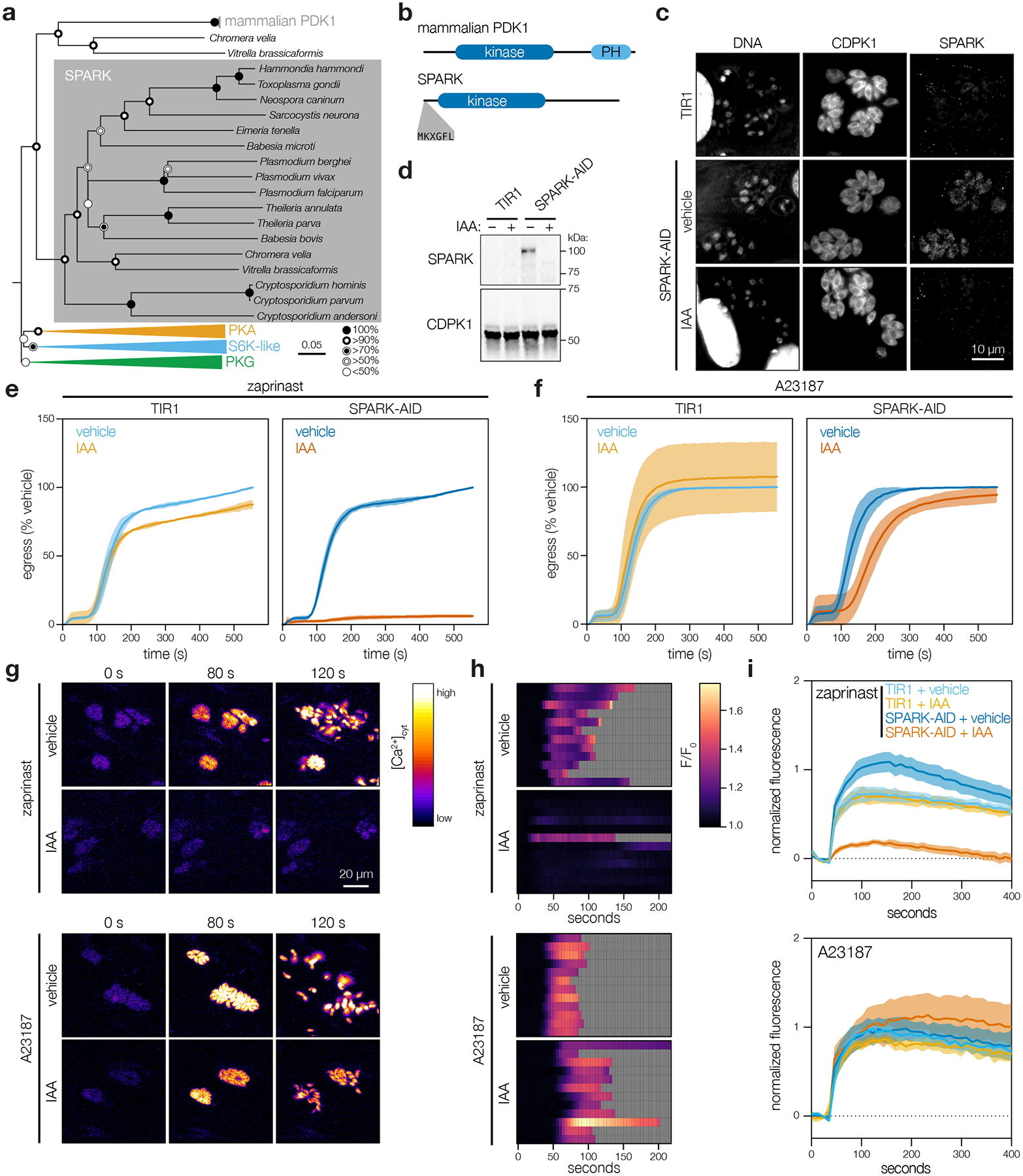
Protein kinases regulate fundamental aspects of eukaryotic cell biology, making them attractive chemotherapeutic targets in parasites like Plasmodium spp. and Toxoplasma gondii. To systematically examine the parasite kinome, we developed a high-throughput tagging (HiT) strategy to endogenously label protein kinases with an auxin-inducible degron and fluorophore. Hundreds of tagging vectors were assembled from synthetic sequences in a single reaction and used to generate pools of mutants to determine localization and function. Examining 1,160 arrayed clones, we assigned 40 protein localizations and associated 15 kinases with distinct defects. The fitness of tagged alleles was also measured by pooled screening, distinguishing delayed from acute phenotypes. A previously unstudied kinase, associated with a delayed phenotype, was shown to be a regulator of invasion and egress. We named the kinase Store Potentiating/Activating Regulatory Kinase (SPARK), based on its impact on intracellular Ca2+ stores. Despite homology to mammalian 3-phosphoinositide-dependent protein kinase-1 (PDK1), SPARK lacks a lipid-binding domain, suggesting a rewiring of the pathway in parasites. HiT screening extends genome-wide approaches into complex cellular phenotypes, providing a scalable and versatile platform to dissect parasite biology.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
COMPETING INTERESTS STATEMENT
The authors declare no competing interests.
Figures
Comment in
-
A CRISPR upgrade unlocks Toxoplasma gene function.Trends Parasitol. 2022 Oct;38(10):826-828. doi: 10.1016/j.pt.2022.07.007. Epub 2022 Aug 13. Trends Parasitol. 2022. PMID: 35973902
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
