

Kinase-independent synthesis of 3-phosphorylated phosphoinositides by a phosphotransferase
- PMID: 35484249
- PMCID: PMC9107517
- DOI: 10.1038/s41556-022-00895-y
Kinase-independent synthesis of 3-phosphorylated phosphoinositides by a phosphotransferase
Abstract
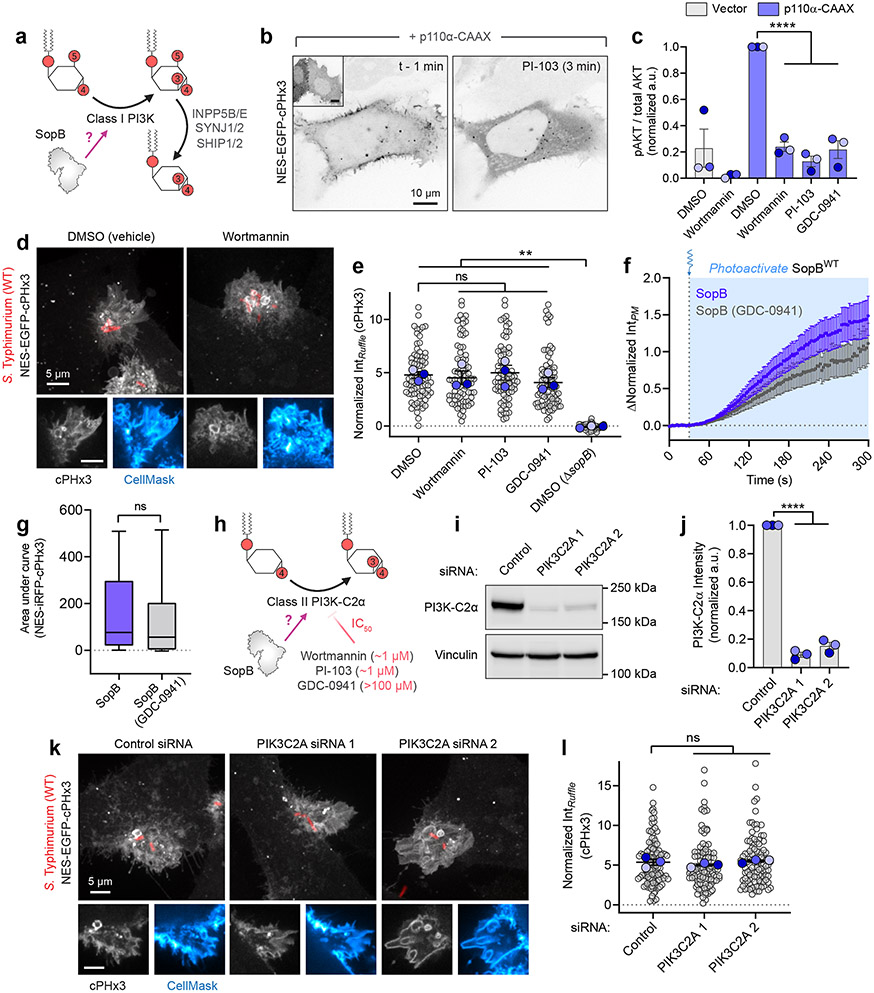
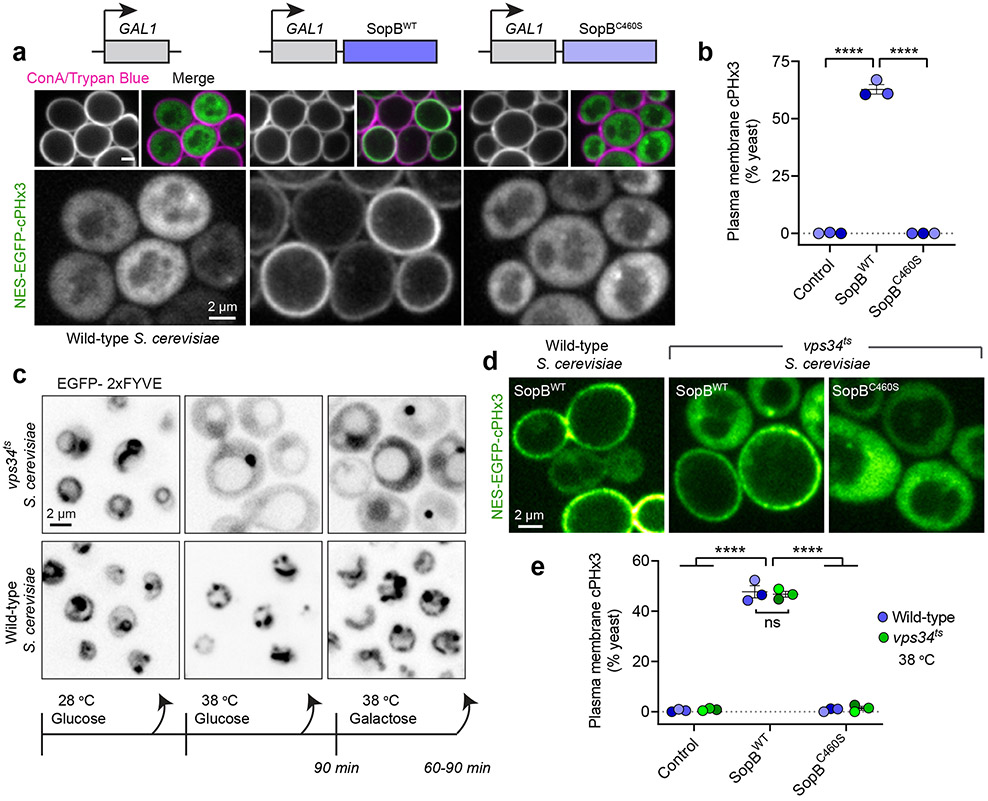
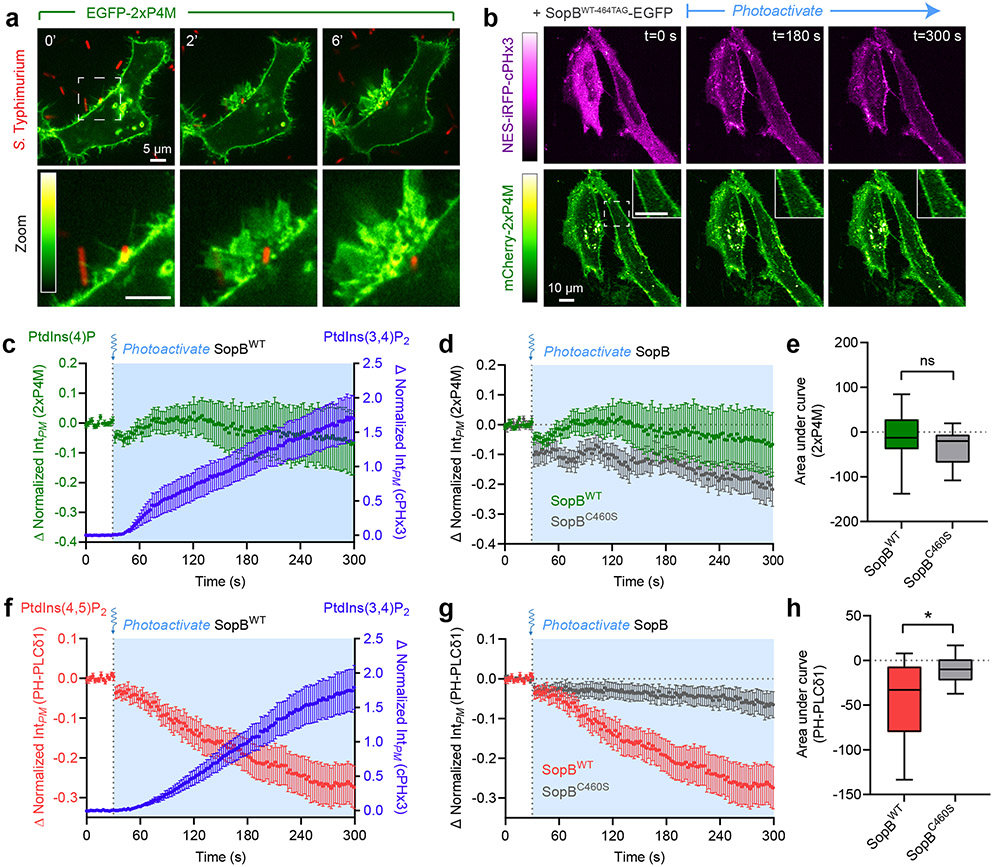
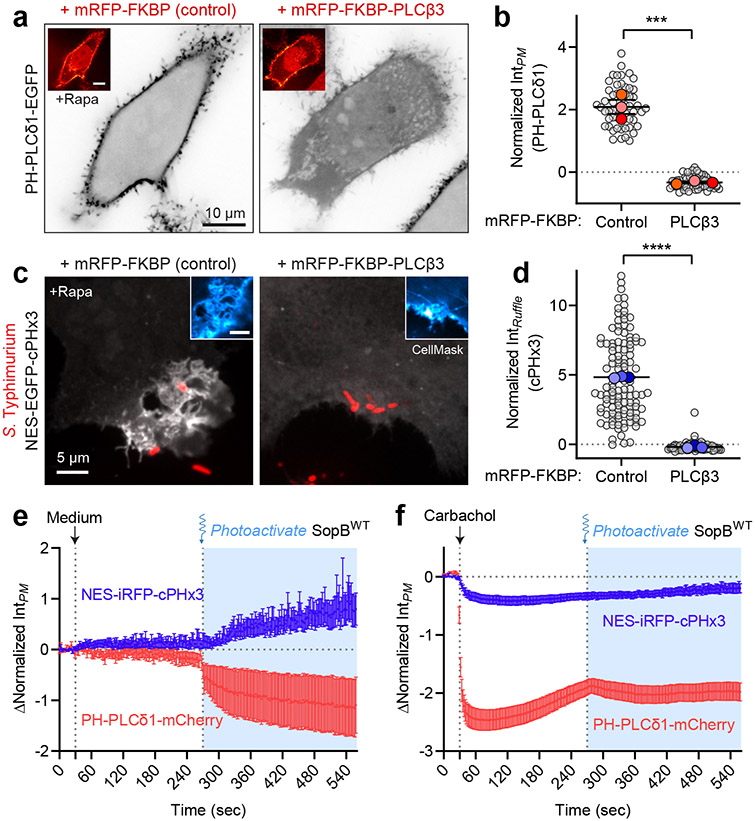
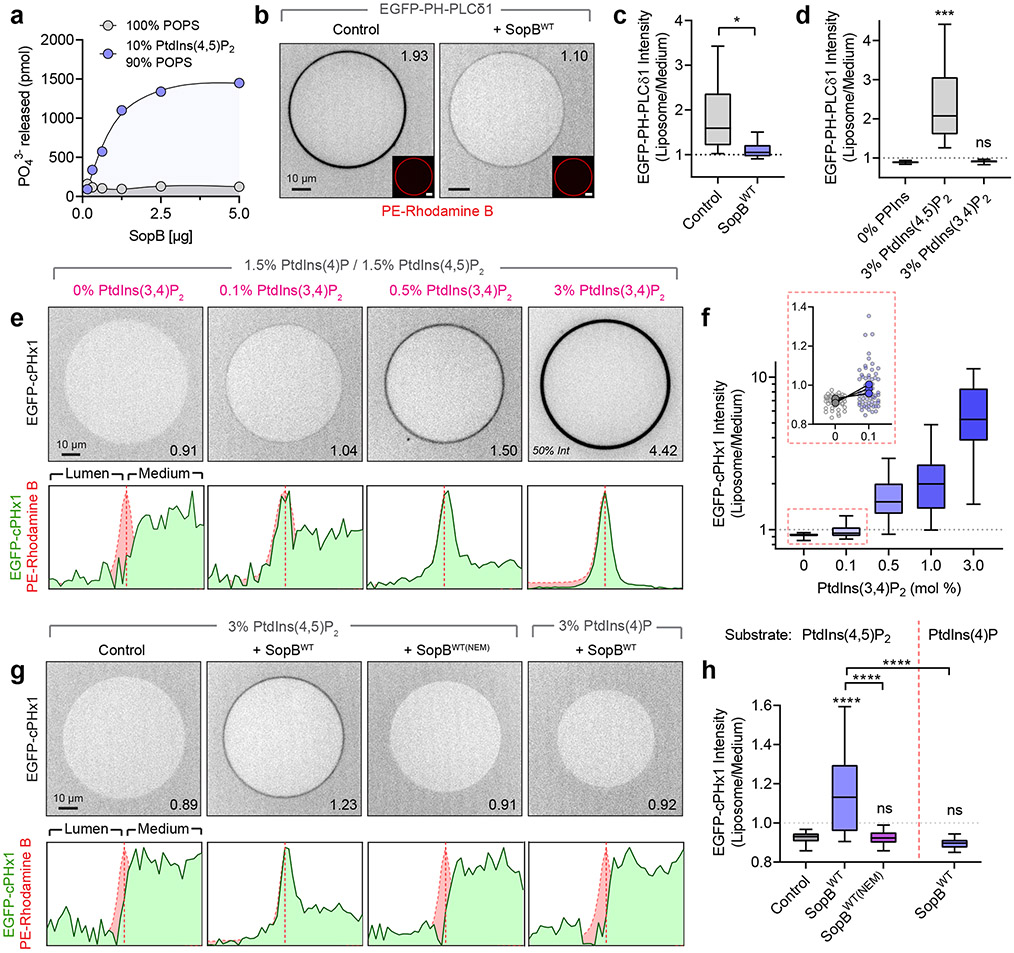
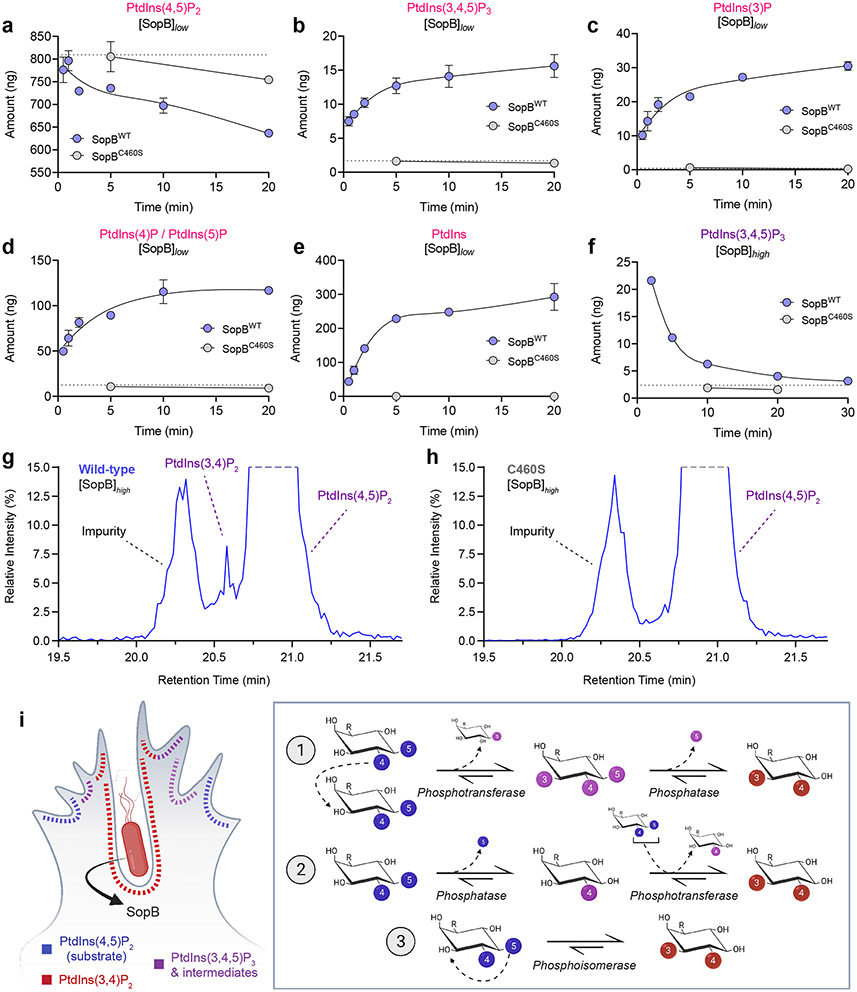
Despite their low abundance, phosphoinositides play a central role in membrane traffic and signalling. PtdIns(3,4,5)P3 and PtdIns(3,4)P2 are uniquely important, as they promote cell growth, survival and migration. Pathogenic organisms have developed means to subvert phosphoinositide metabolism to promote successful infection and their survival in host organisms. We demonstrate that PtdIns(3,4)P2 is a major product generated in host cells by the effectors of the enteropathogenic bacteria Salmonella and Shigella. Pharmacological, gene silencing and heterologous expression experiments revealed that, remarkably, the biosynthesis of PtdIns(3,4)P2 occurs independently of phosphoinositide 3-kinases. Instead, we found that the Salmonella effector SopB, heretofore believed to be a phosphatase, generates PtdIns(3,4)P2 de novo via a phosphotransferase/phosphoisomerase mechanism. Recombinant SopB is capable of generating PtdIns(3,4,5)P3 and PtdIns(3,4)P2 from PtdIns(4,5)P2 in a cell-free system. Through a remarkable instance of convergent evolution, bacterial effectors acquired the ability to synthesize 3-phosphorylated phosphoinositides by an ATP- and kinase-independent mechanism, thereby subverting host signalling to gain entry and even provoke oncogenic transformation.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Figures
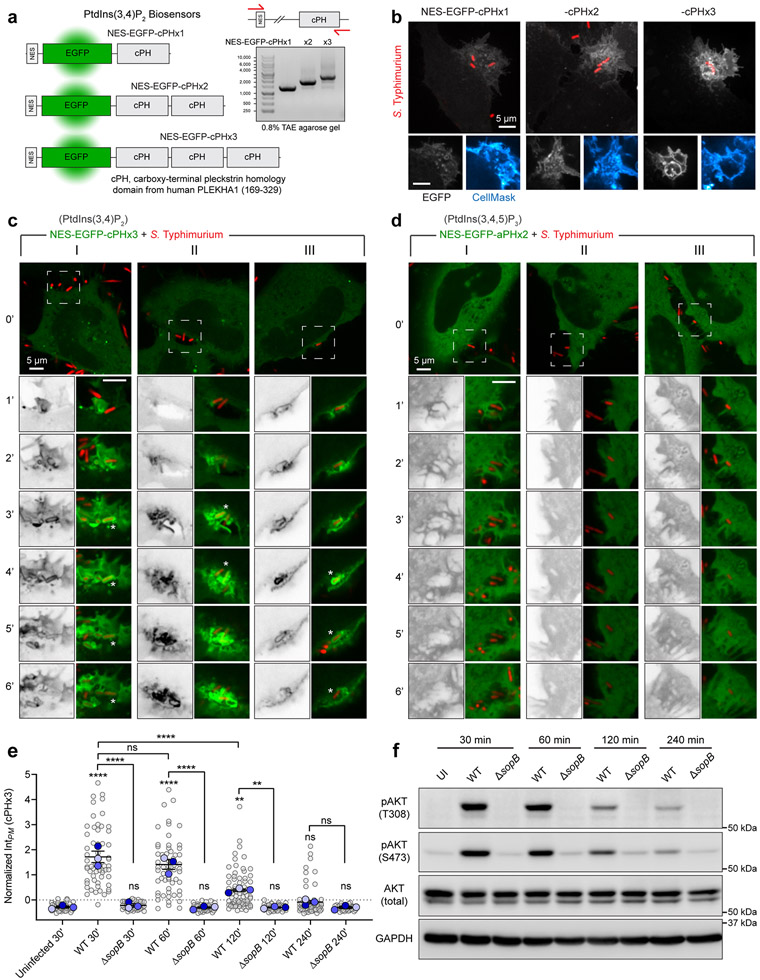
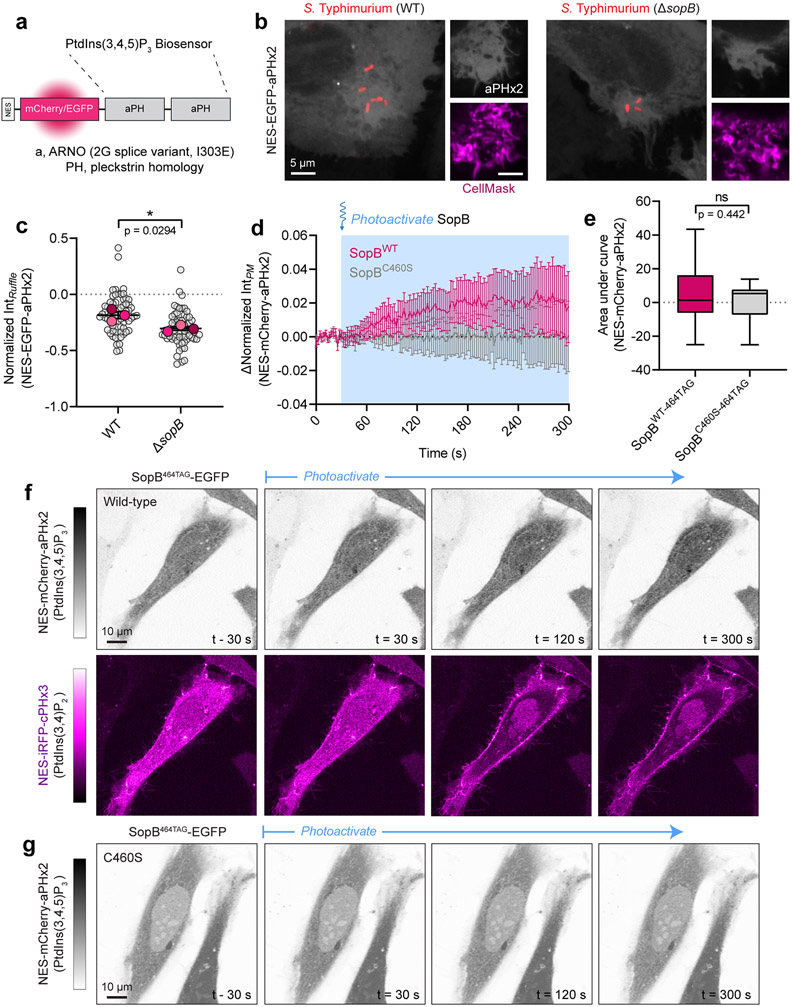

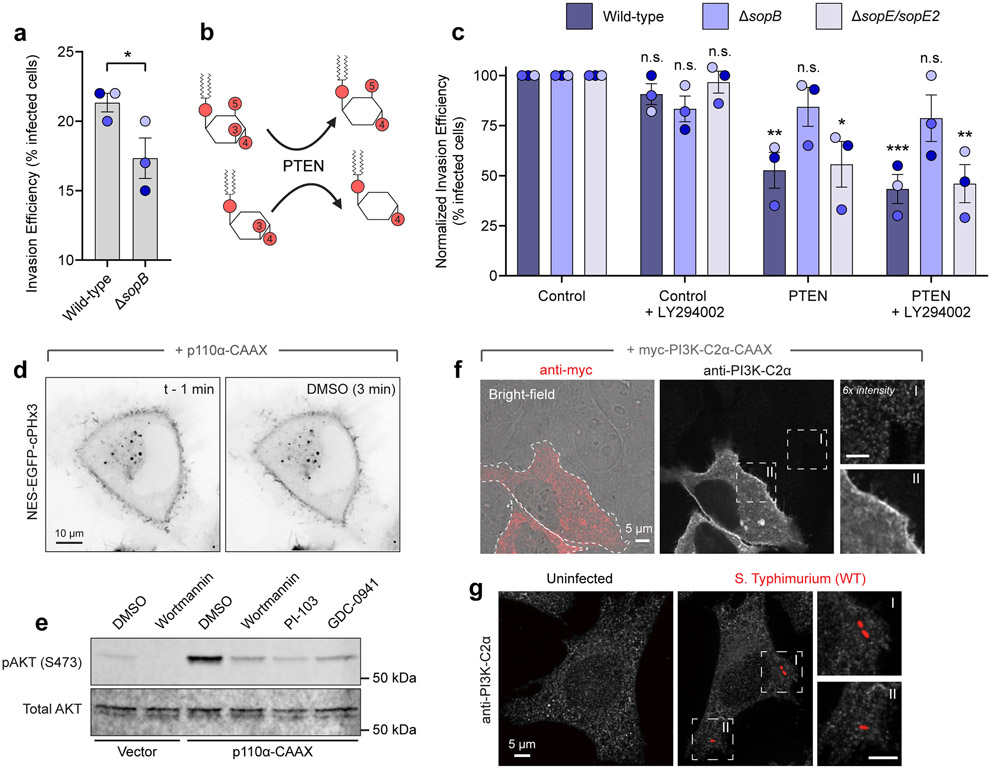
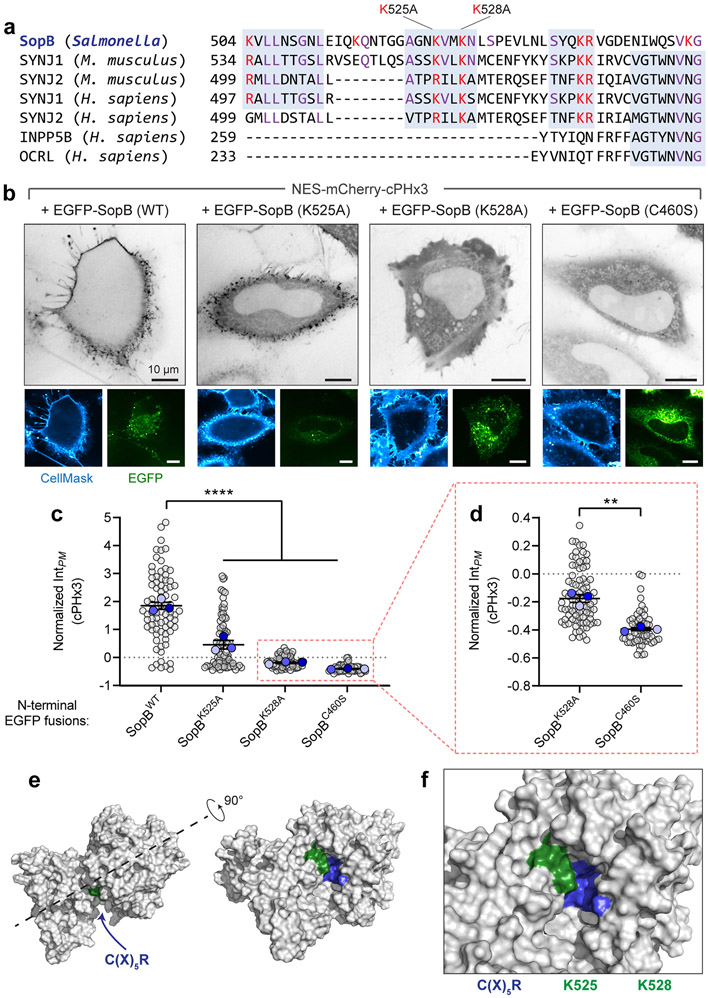
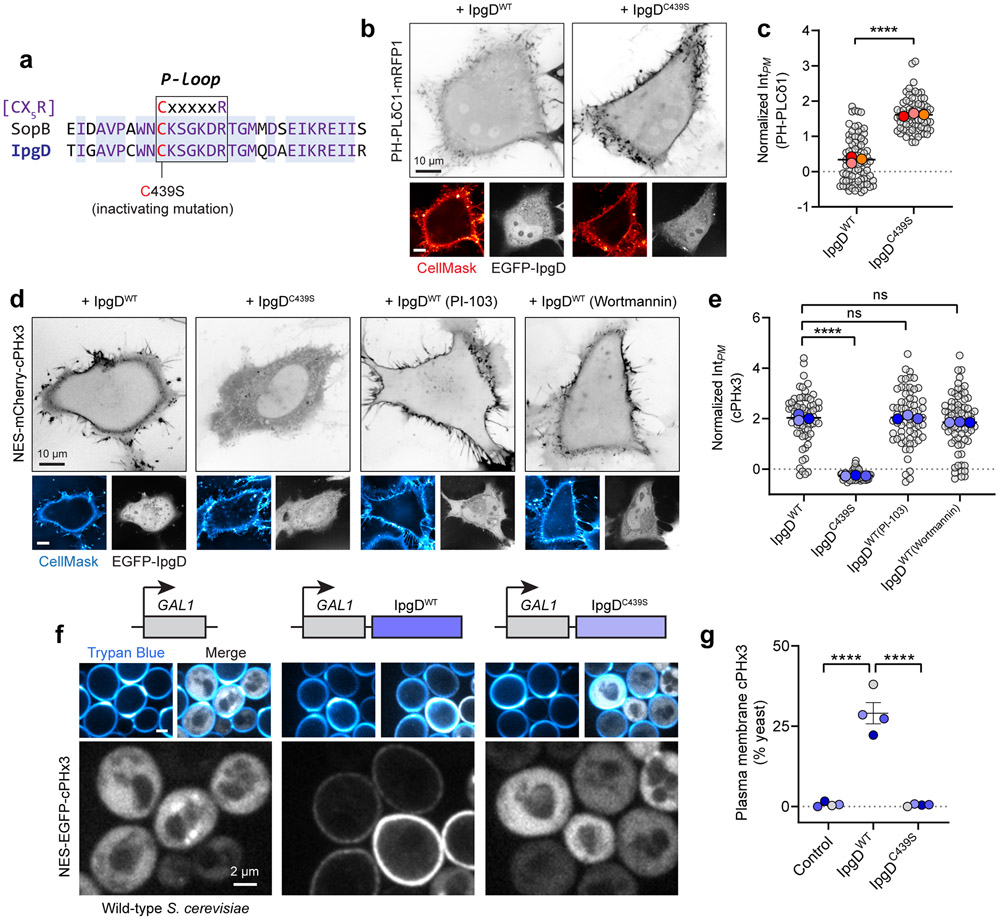
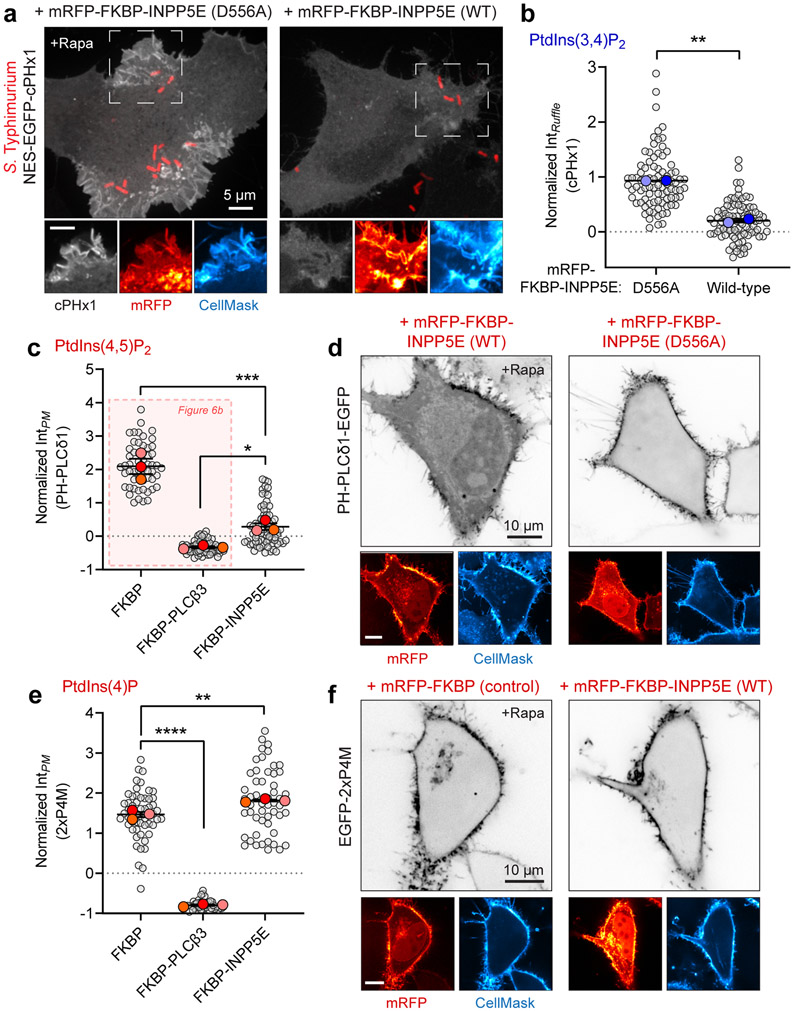

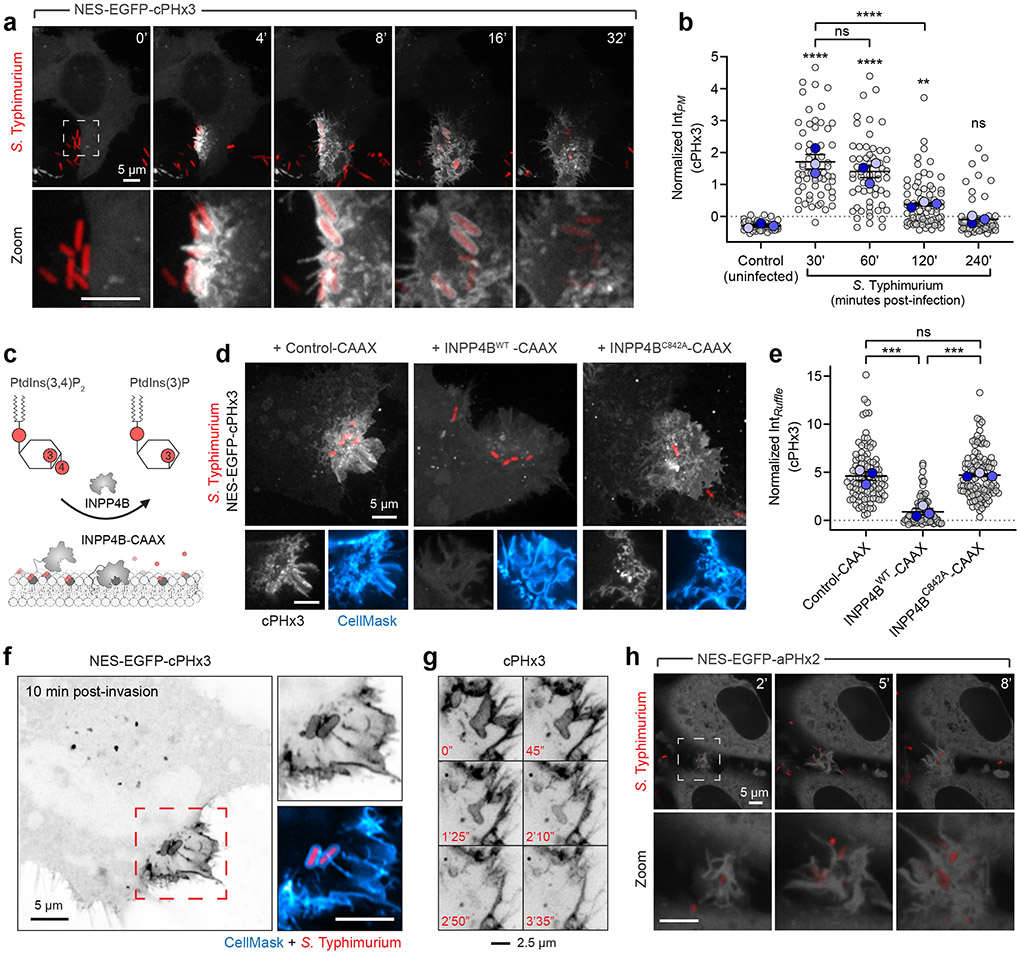
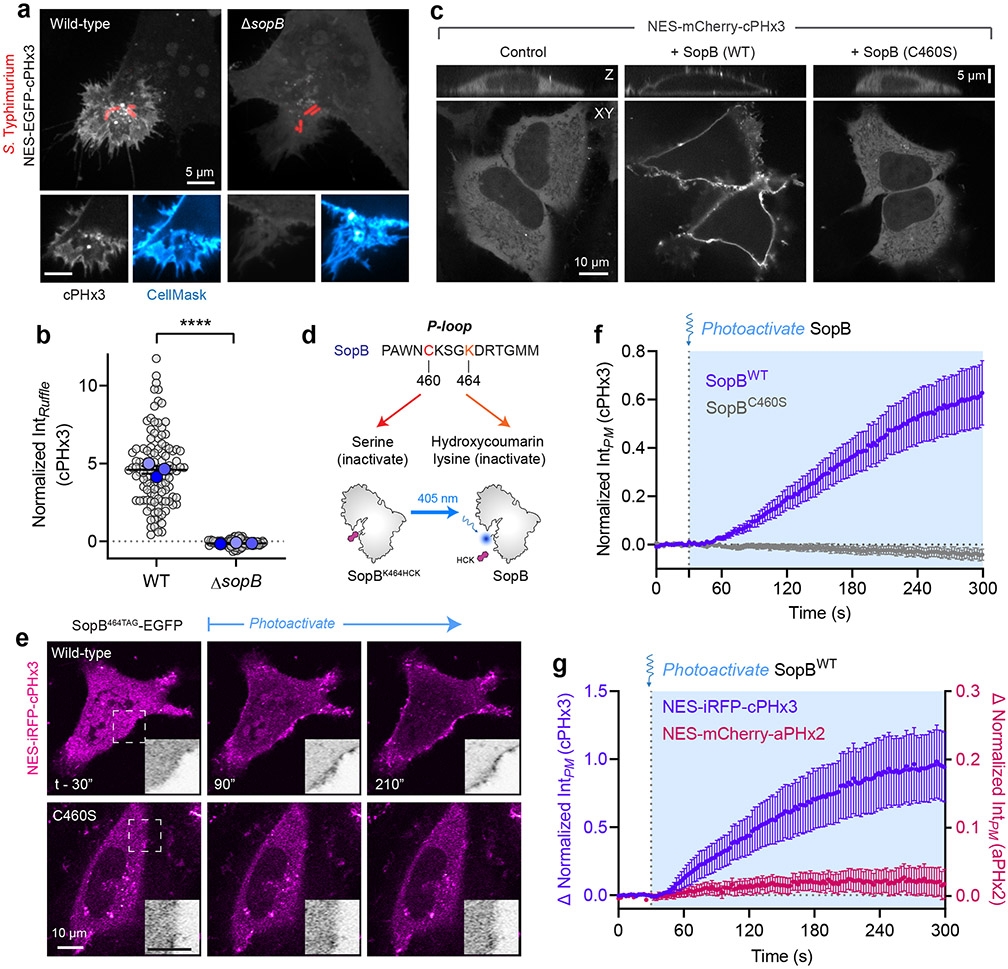
Comment in
-
Phosphoinositide phosphorylation sans kinase.Nat Cell Biol. 2022 May;24(5):604-606. doi: 10.1038/s41556-022-00885-0. Nat Cell Biol. 2022. PMID: 35484248 No abstract available.
References
-
- Hakim S, Bertucci MC, Conduit SE, Vuong DL & Mitchell CA Inositol polyphosphate phosphatases in human disease. Current topics in microbiology and immunology 362, 247–314 (2012). - PubMed
-
- Sasaki T et al. Mammalian phosphoinositide kinases and phosphatases. Progress in Lipid Research 48, 307–343 (2009). - PubMed
-
- Stephens LR, Jackson TR & Hawkins PT Agonist-stimulated synthesis of phosphatidylinositol(3,4,5)-trisphosphate: a new intracellular signalling system? Biochimica et biophysica acta 1179, 27–75 (1993). - PubMed
-
- Bilanges B, Posor Y & Vanhaesebroeck B PI3K isoforms in cell signalling and vesicle trafficking. Nature Reviews Molecular Cell Biology 20, 515–534 (2019). - PubMed
Methods References
-
- Leibiger B et al. Insulin-feedback via PI3K-C2alpha activated PKBalpha/Akt1 is required for glucose-stimulated insulin secretion. FASEB journal : official publication of the Federation of American Societies for Experimental Biology 24, 1824–37 (2010). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
