

Why succinate? Physiological regulation by a mitochondrial coenzyme Q sentinel
- PMID: 35484255
- PMCID: PMC9150600
- DOI: 10.1038/s41589-022-01004-8
Why succinate? Physiological regulation by a mitochondrial coenzyme Q sentinel
Abstract
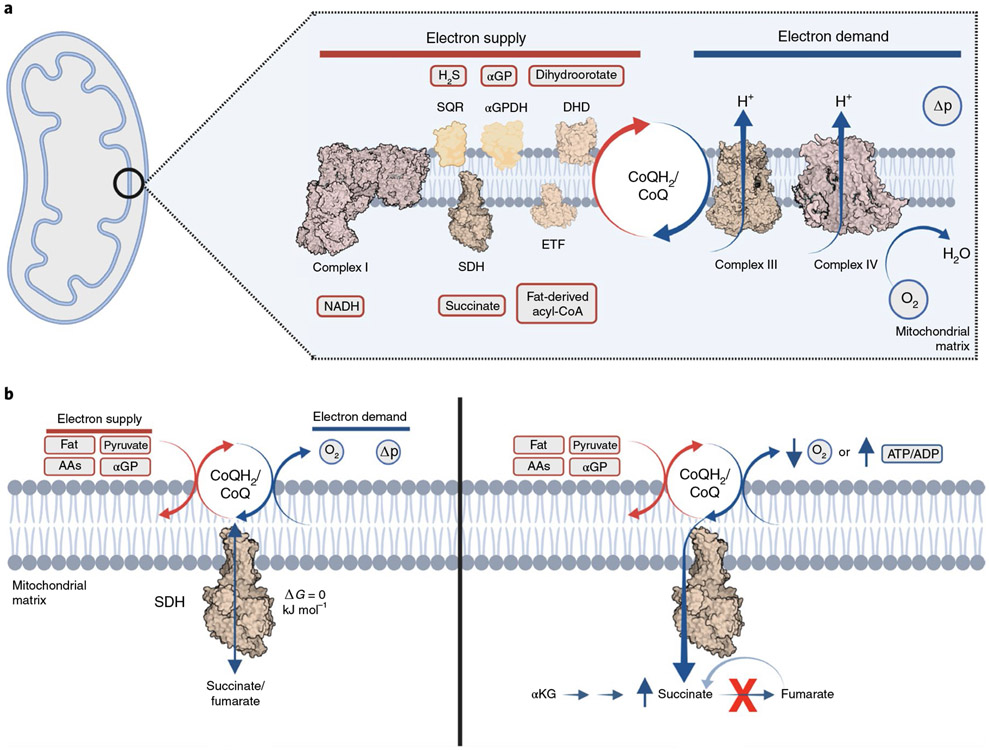
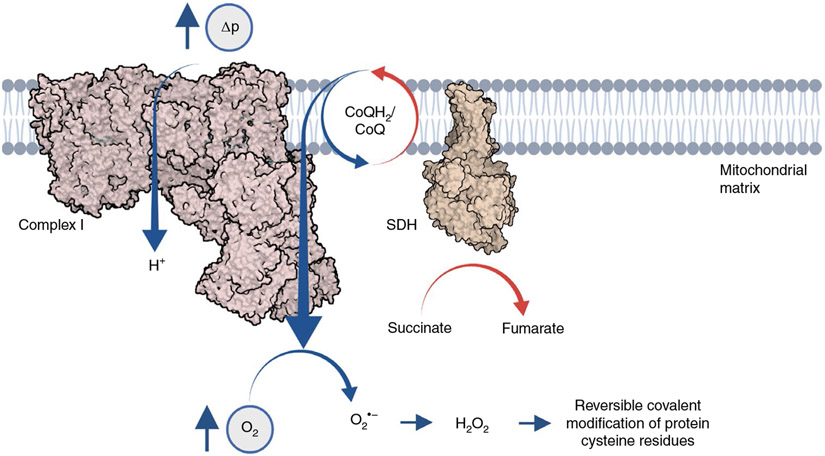
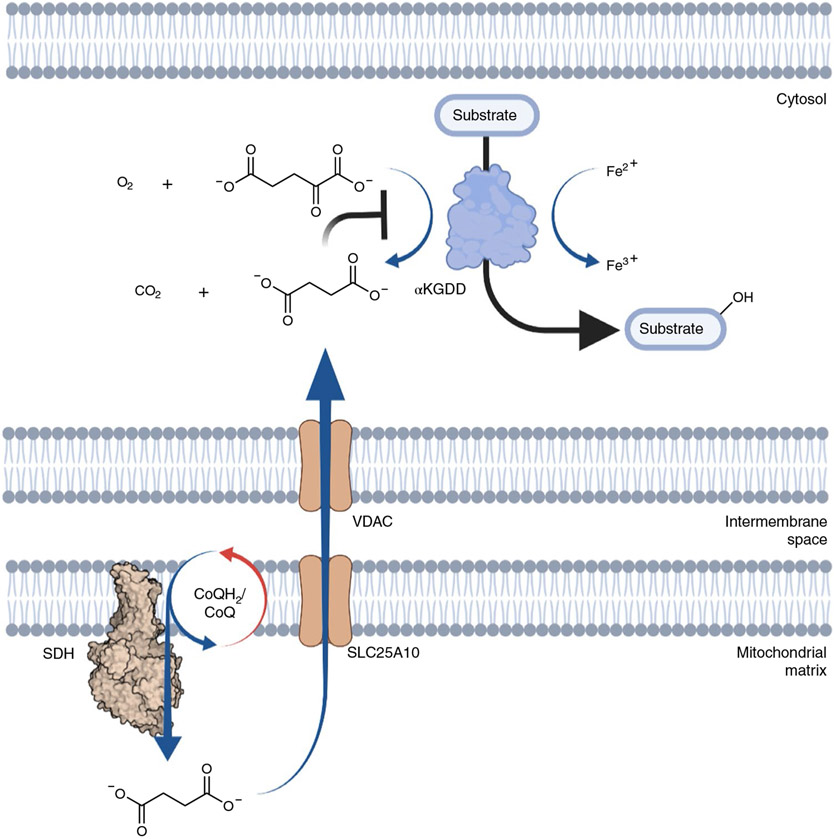
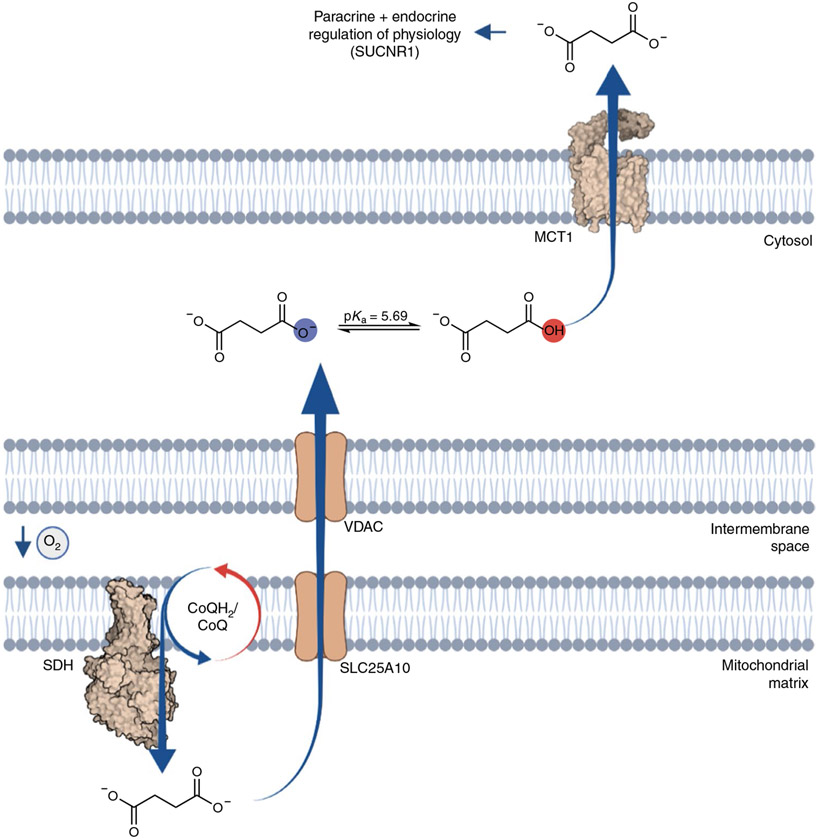
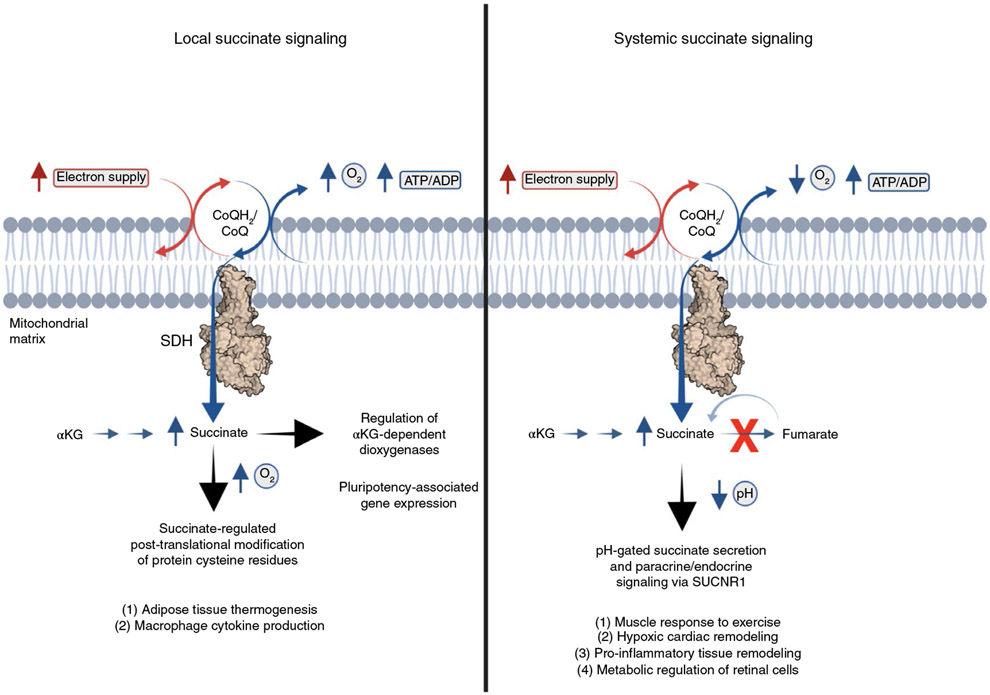
Metabolites once considered solely in catabolism or anabolism turn out to have key regulatory functions. Among these, the citric acid cycle intermediate succinate stands out owing to its multiple roles in disparate pathways, its dramatic concentration changes and its selective cell release. Here we propose that succinate has evolved as a signaling modality because its concentration reflects the coenzyme Q (CoQ) pool redox state, a central redox couple confined to the mitochondrial inner membrane. This connection is of general importance because CoQ redox state integrates three bioenergetic parameters: mitochondrial electron supply, oxygen tension and ATP demand. Succinate, by equilibrating with the CoQ pool, enables the status of this central bioenergetic parameter to be communicated from mitochondria to the rest of the cell, into the circulation and to other cells. The logic of this form of regulation explains many emerging roles of succinate in biology, and suggests future research questions.
© 2022. Springer Nature America, Inc.
Figures
References
-
- Murphy MP & O’Neill LAJ Krebs cycle reimagined: the emerging roles of succinate and itaconate as signal transducers. Cell 174, 780–784 (2018). - PubMed
-
- Winther S, Trauelsen M & Schwartz TW Protective succinate-SUCNR1 metabolic stress signaling gone bad. Cell Metab. 33, 1276–1278 (2021). - PubMed
-
- Chandel NS Evolution of mitochondria as signaling organelles. Cell Metab. 22, 204–206 (2015). - PubMed
-
- Nicholls DG & Ferguson SJ Bioenergetics 3rd edn, 31–55 (Academic Press, 2003).
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
