

Spindle-slow oscillation coupling correlates with memory performance and connectivity changes in a hippocampal network after sleep
- PMID: 35488512
- PMCID: PMC9374888
- DOI: 10.1002/hbm.25893
Spindle-slow oscillation coupling correlates with memory performance and connectivity changes in a hippocampal network after sleep
Abstract
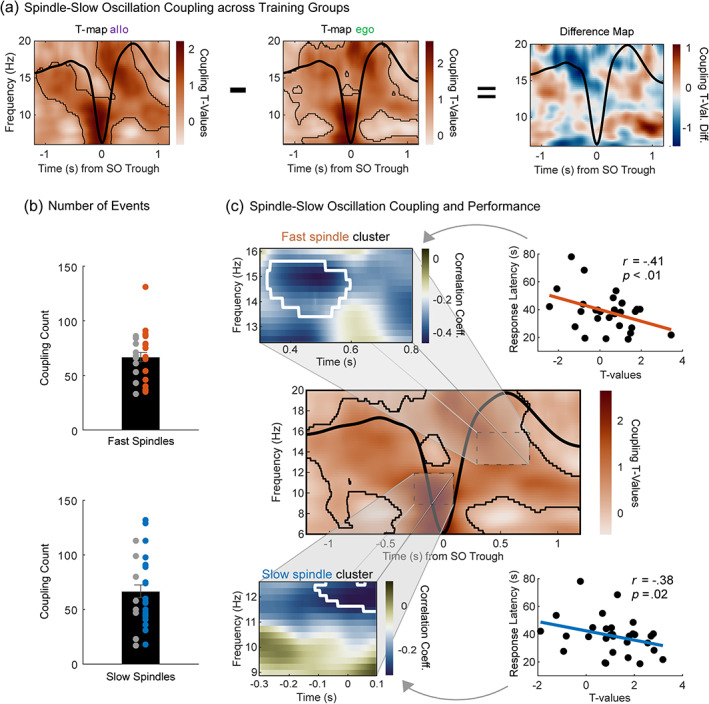
After experiences are encoded, post-encoding reactivations during sleep have been proposed to mediate long-term memory consolidation. Spindle-slow oscillation coupling during NREM sleep is a candidate mechanism through which a hippocampal-cortical dialogue may strengthen a newly formed memory engram. Here, we investigated the role of fast spindle- and slow spindle-slow oscillation coupling in the consolidation of spatial memory in humans with a virtual watermaze task involving allocentric and egocentric learning strategies. Furthermore, we analyzed how resting-state functional connectivity evolved across learning, consolidation, and retrieval of this task using a data-driven approach. Our results show task-related connectivity changes in the executive control network, the default mode network, and the hippocampal network at post-task rest. The hippocampal network could further be divided into two subnetworks of which only one showed modulation by sleep. Decreased functional connectivity in this subnetwork was associated with higher spindle-slow oscillation coupling power, which was also related to better memory performance at test. Overall, this study contributes to a more holistic understanding of the functional resting-state networks and the mechanisms during sleep associated to spatial memory consolidation.
Keywords: memory consolidation; resting-state networks; sleep; slow oscillations; spindles.
© 2022 The Authors. Human Brain Mapping published by Wiley Periodicals LLC.
Figures




References
-
- Akaike, H. (1973). Maximum likelihood identification of Gaussian autoregressive moving average models. Biometrika, 60(2), 255–265.
-
- Albouy, G. , Fogel, S. , King, B. R. , Laventure, S. , Benali, H. , Karni, A. , Carrier, J. , Robertson, E. M. , & Doyon, J. (2015). Maintaining vs. enhancing motor sequence memories: Respective roles of striatal and hippocampal systems. NeuroImage, 108, 423–434. 10.1016/j.neuroimage.2014.12.049 - DOI - PubMed
-
- Anderer, P. , Klösch, G. , Gruber, G. , Trenker, E. , Pascual‐Marqui, R. D. , Zeitlhofer, J. , Barbanoj, M. J. , Rappelsberger, P. , & Saletu, B. (2001). Low‐resolution brain electromagnetic tomography revealed simultaneously active frontal and parietal sleep spindle sources in the human cortex. Neuroscience, 103(3), 581–592. - PubMed
-
- Barakat, M. , Doyon, J. , Debas, K. , Vandewalle, G. , Morin, A. , Poirier, G. , Martin, N. , Lafortune, M. , Karni, A. , Ungerleider, L. G. , Benali, H. , & Carrier, J. (2011). Fast and slow spindle involvement in the consolidation of a new motor sequence. Behavioural Brain Research, 217(1), 117–121. 10.1016/j.bbr.2010.10.019 - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
