

β-Catenin Sustains and Is Required for YES-associated Protein Oncogenic Activity in Cholangiocarcinoma
- PMID: 35489428
- PMCID: PMC9329198
- DOI: 10.1053/j.gastro.2022.04.028
β-Catenin Sustains and Is Required for YES-associated Protein Oncogenic Activity in Cholangiocarcinoma
Abstract
Background & aims: YES-associated protein (YAP) aberrant activation is implicated in intrahepatic cholangiocarcinoma (iCCA). Transcriptional enhanced associate domain (TEAD)-mediated transcriptional regulation is the primary signaling event downstream of YAP. The role of Wnt/β-Catenin signaling in cholangiocarcinogenesis remains undetermined. Here, we investigated the possible molecular interplay between YAP and β-Catenin cascades in iCCA.
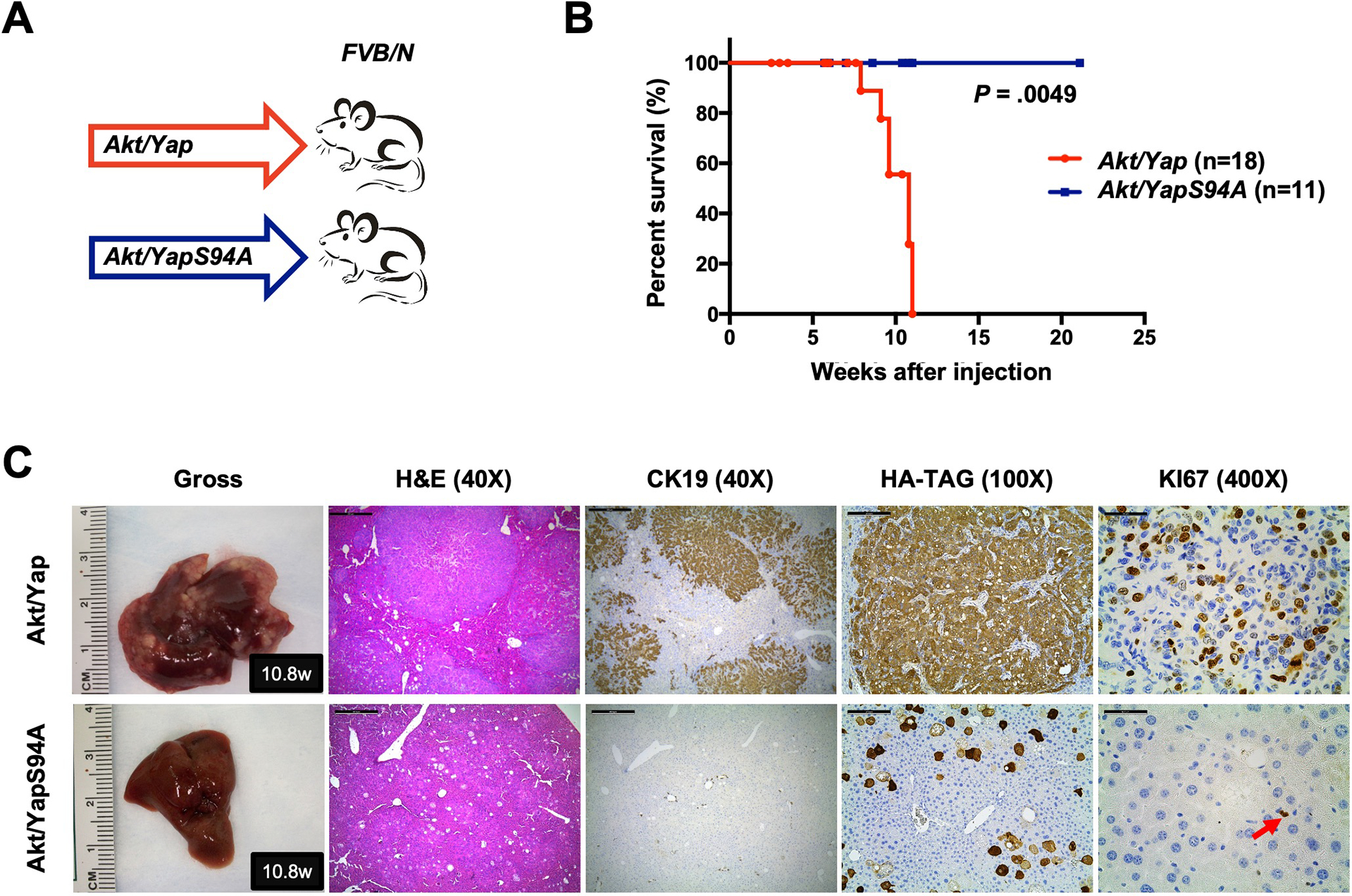
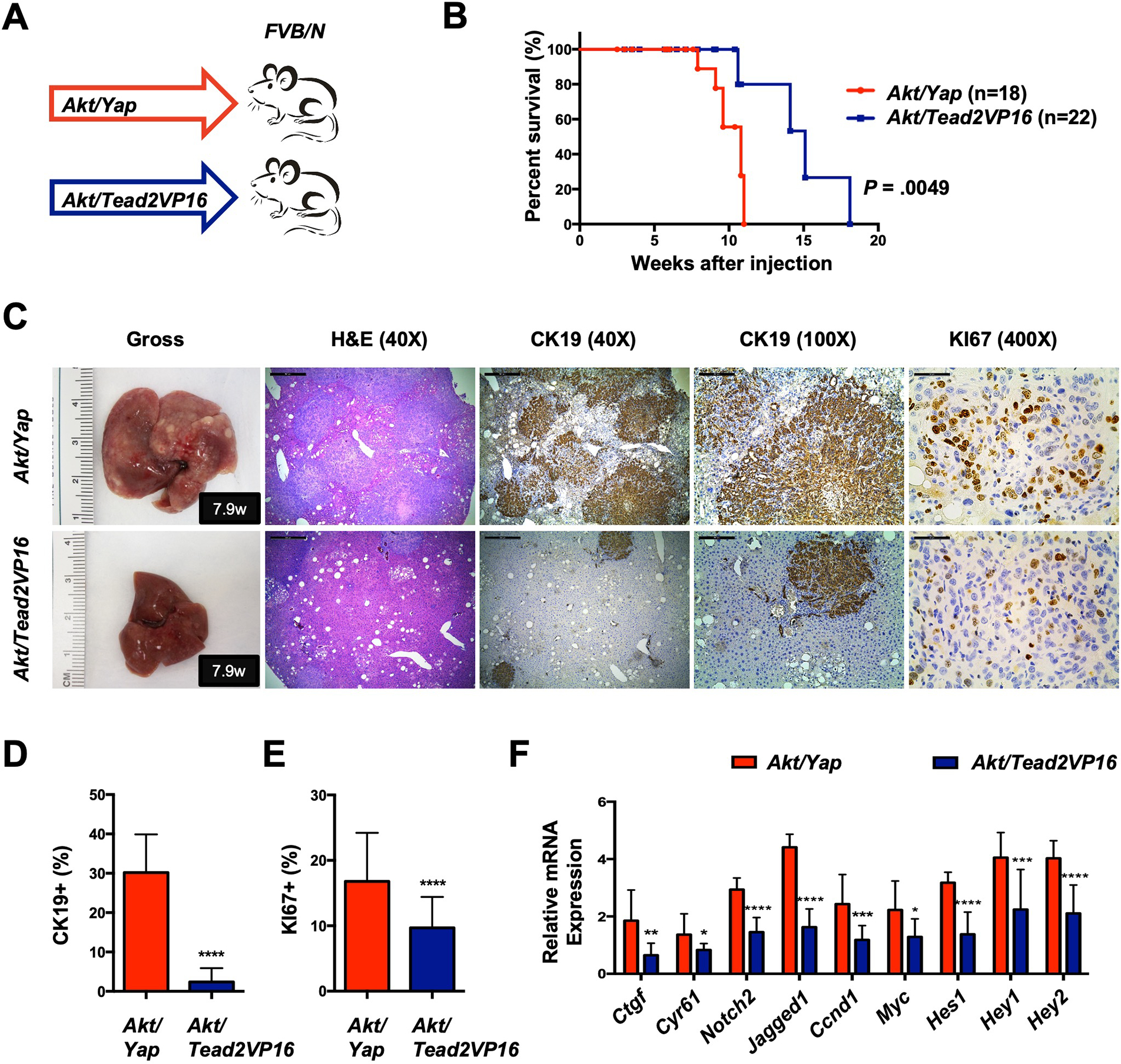
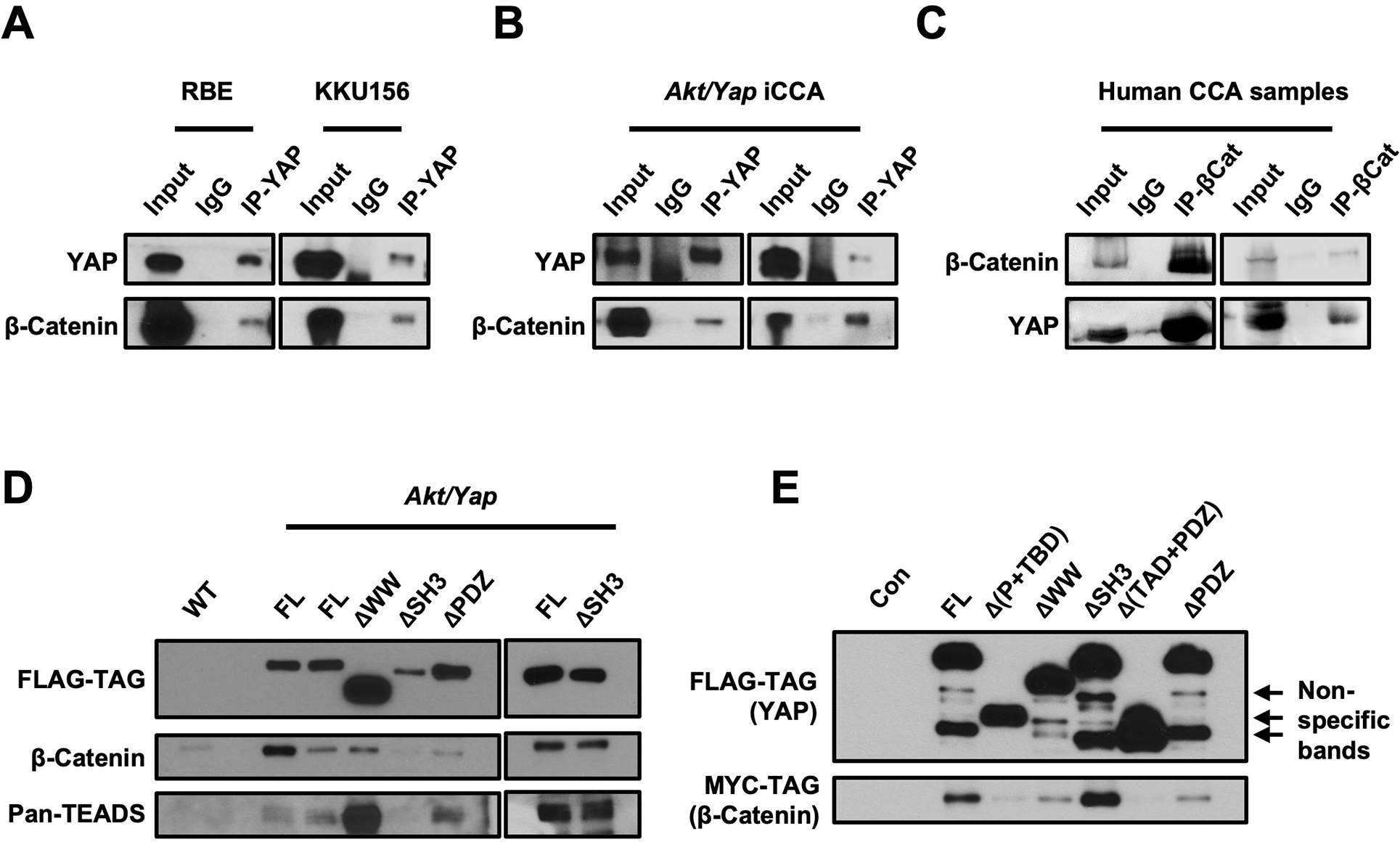
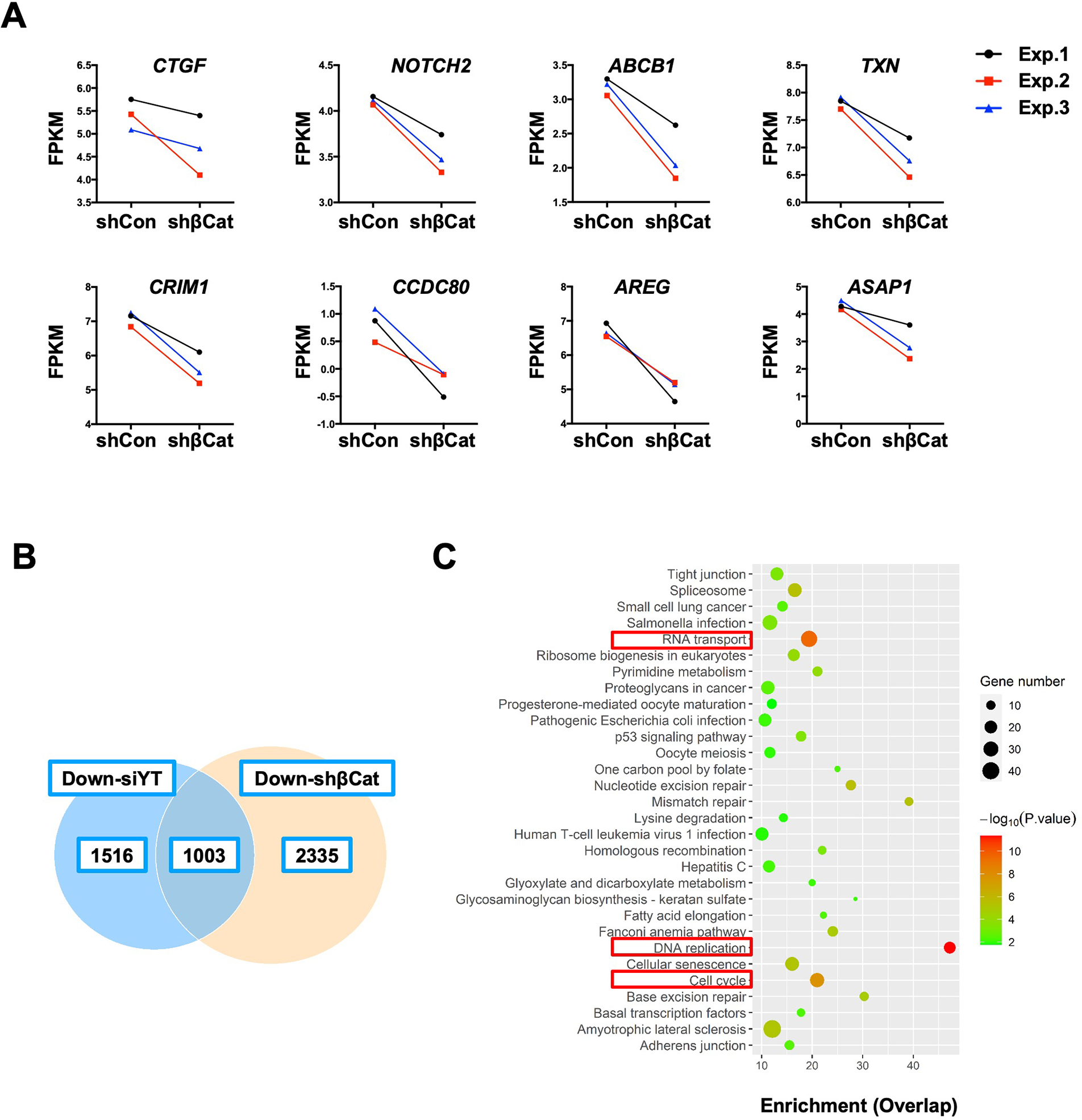
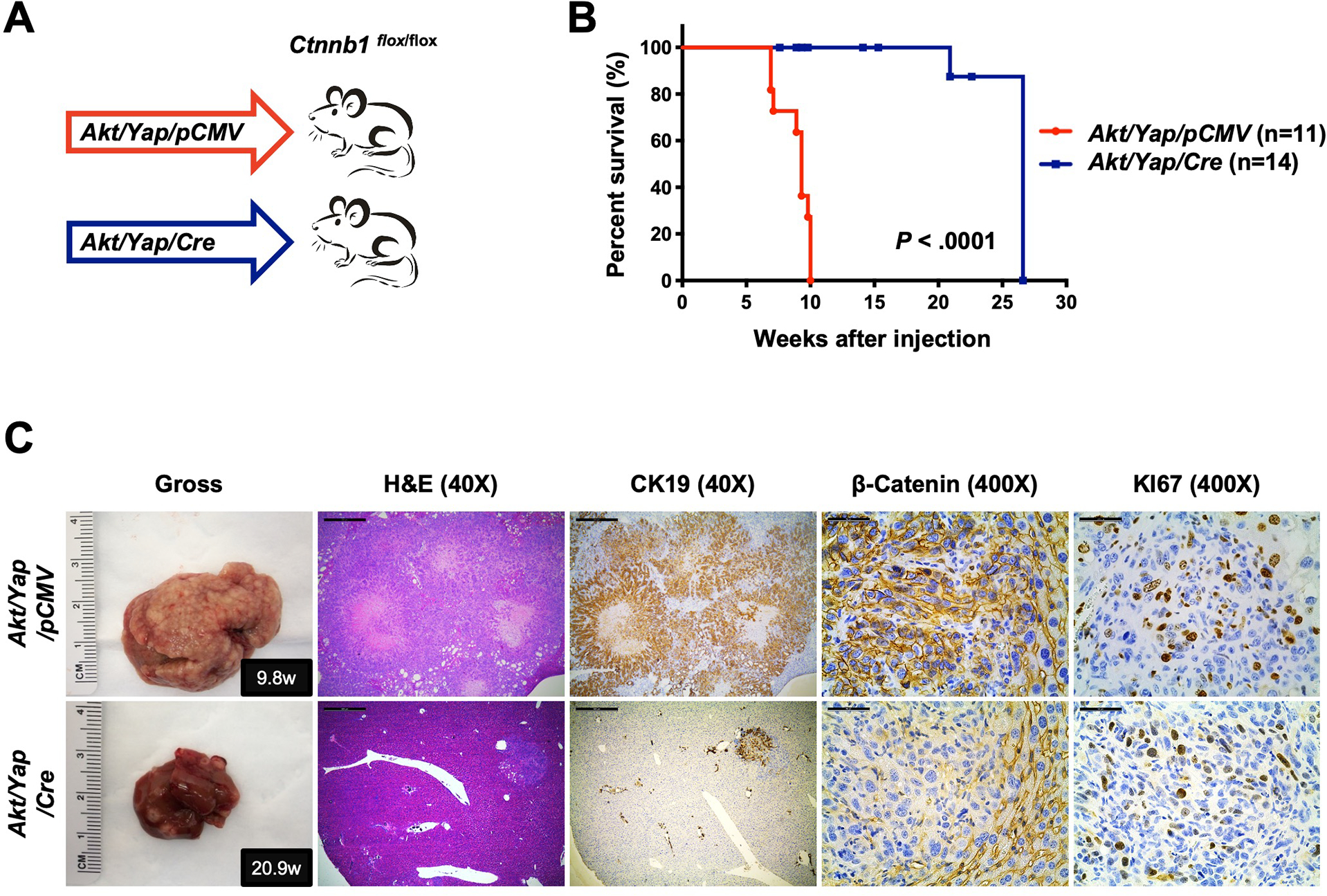
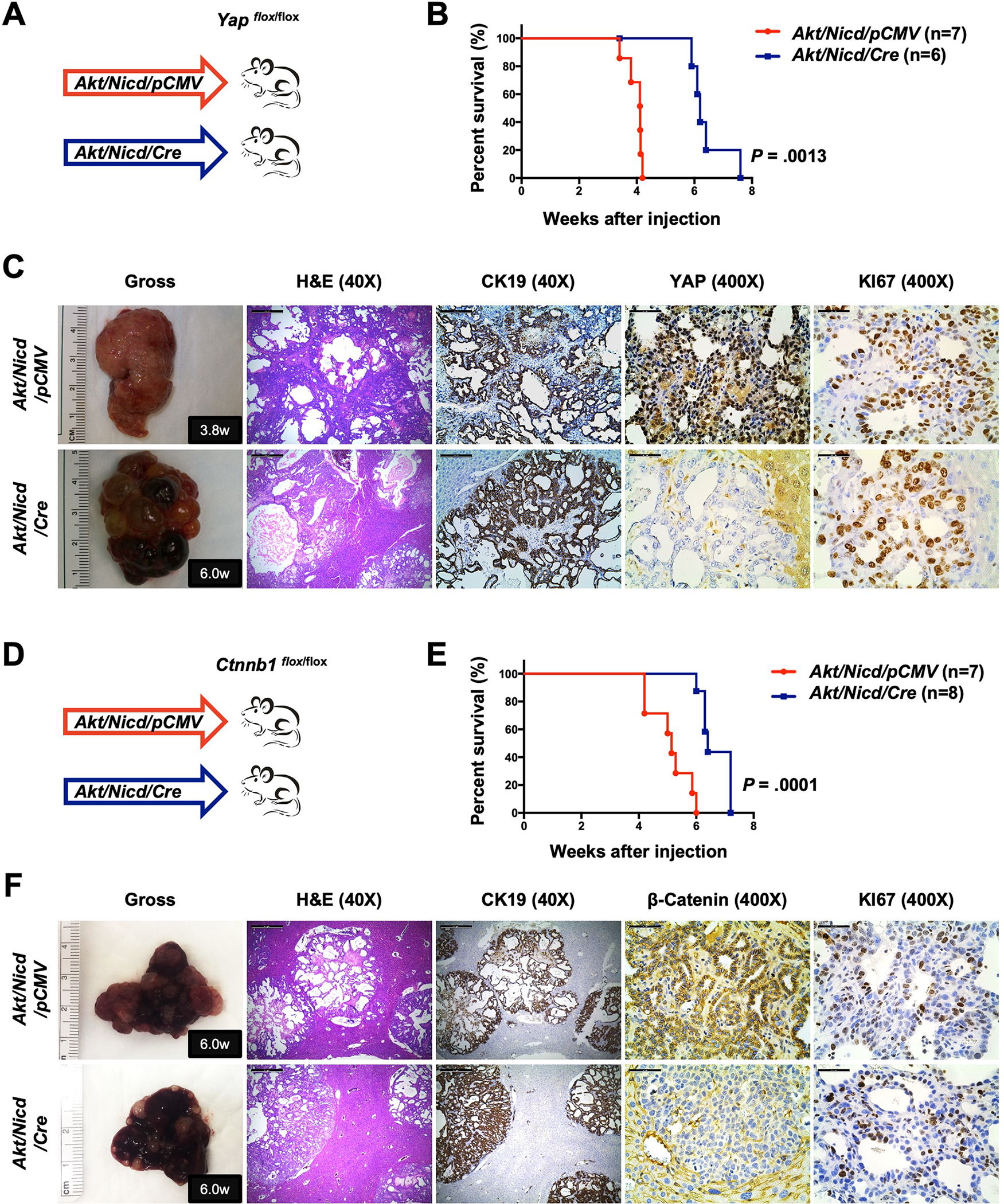
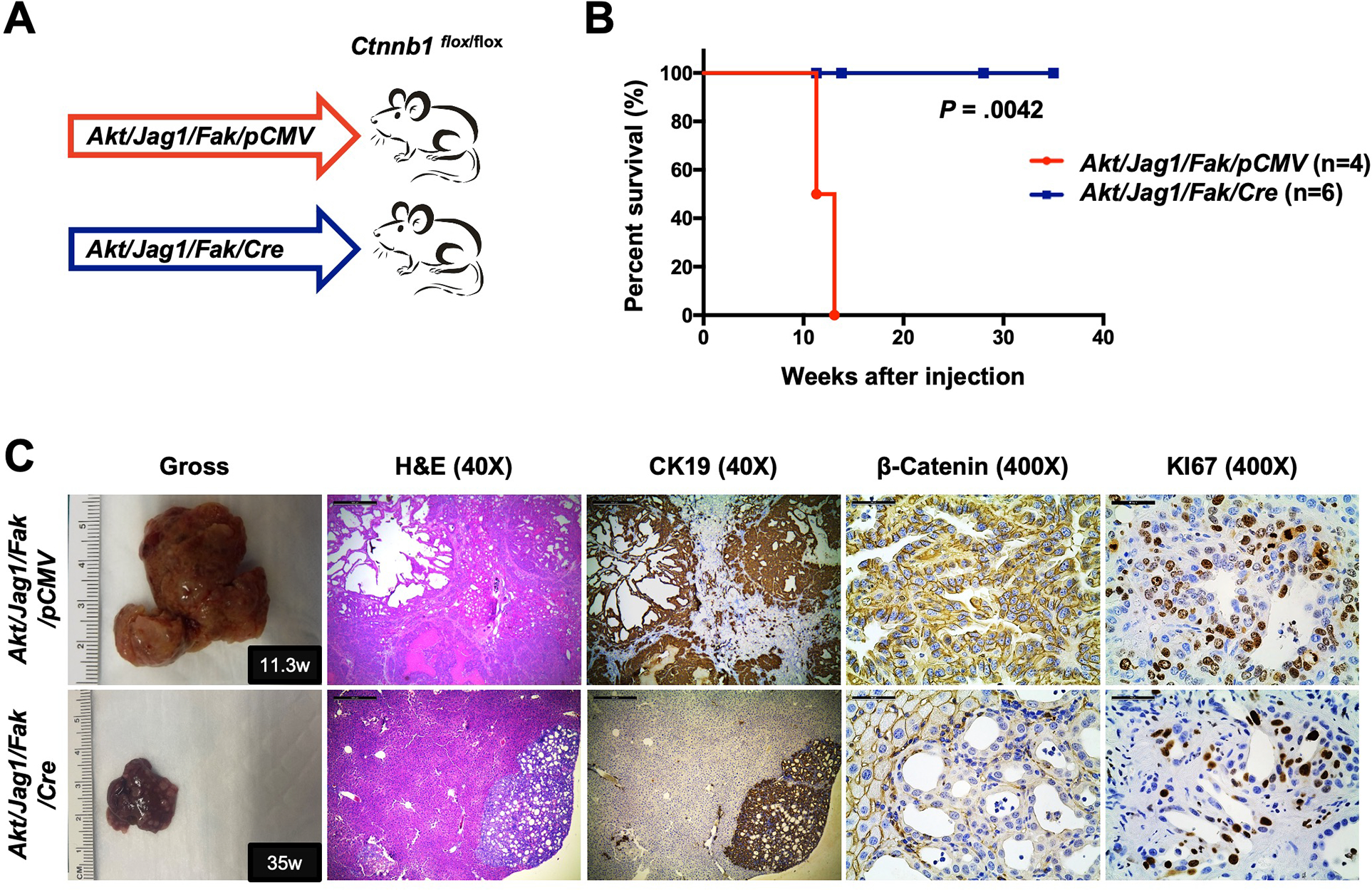
Methods: Activated AKT (Myr-Akt) was coexpressed with YAP (YapS127A) or Tead2VP16 via hydrodynamic tail vein injection into mouse livers. Tumor growth was monitored, and liver tissues were collected and analyzed using histopathologic and molecular analysis. YAP, β-Catenin, and TEAD interaction in iCCAs was investigated through coimmunoprecipitation. Conditional Ctnnb1 knockout mice were used to determine β-Catenin function in murine iCCA models. RNA sequencing was performed to analyze the genes regulated by YAP and/or β-Catenin. Immunostaining of total and nonphosphorylated/activated β-Catenin staining was performed in mouse and human iCCAs.
Results: We discovered that TEAD factors are required for YAP-dependent iCCA development. However, transcriptional activation of TEADs did not fully recapitulate YAP's activities in promoting cholangiocarcinogenesis. Notably, β-Catenin physically interacted with YAP in human and mouse iCCA. Ctnnb1 ablation strongly suppressed human iCCA cell growth and Yap-dependent cholangiocarcinogenesis. Furthermore, RNA-sequencing analysis revealed that YAP/ transcriptional coactivator with PDZ-binding motif (TAZ) regulate a set of genes significantly overlapping with those controlled by β-Catenin. Importantly, activated/nonphosphorylated β-Catenin was detected in more than 80% of human iCCAs.
Conclusion: YAP induces cholangiocarcinogenesis via TEAD-dependent transcriptional activation and interaction with β-Catenin. β-Catenin binds to YAP in iCCA and is required for YAP full transcriptional activity, revealing the functional crosstalk between YAP and β-Catenin pathways in cholangiocarcinogenesis.
Keywords: Hippo/YAP; Intrahepatic Cholangiocarcinoma; TEADs; β-Catenin.
Copyright © 2022 AGA Institute. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Disclosures
The authors declare no potential conflicts of interest.
Figures
Comment in
-
β-Catenin Is a Novel Target in YES-associated Protein-driven Cholangiocarcinoma.Gastroenterology. 2022 Aug;163(2):374-376. doi: 10.1053/j.gastro.2022.05.047. Epub 2022 Jun 2. Gastroenterology. 2022. PMID: 35661722 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
