

Microbiota-mediated skewing of tryptophan catabolism modulates CD4+ T cells in lupus-prone mice
- PMID: 35494242
- PMCID: PMC9051618
- DOI: 10.1016/j.isci.2022.104241
Microbiota-mediated skewing of tryptophan catabolism modulates CD4+ T cells in lupus-prone mice
Abstract
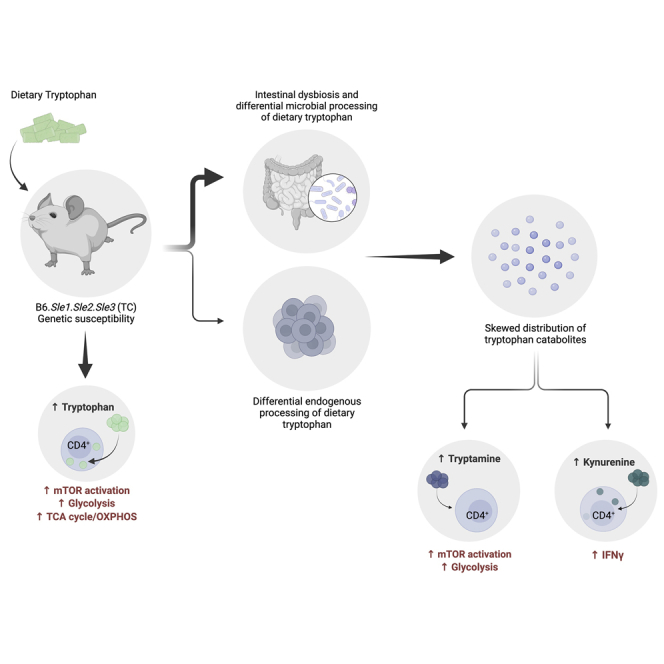
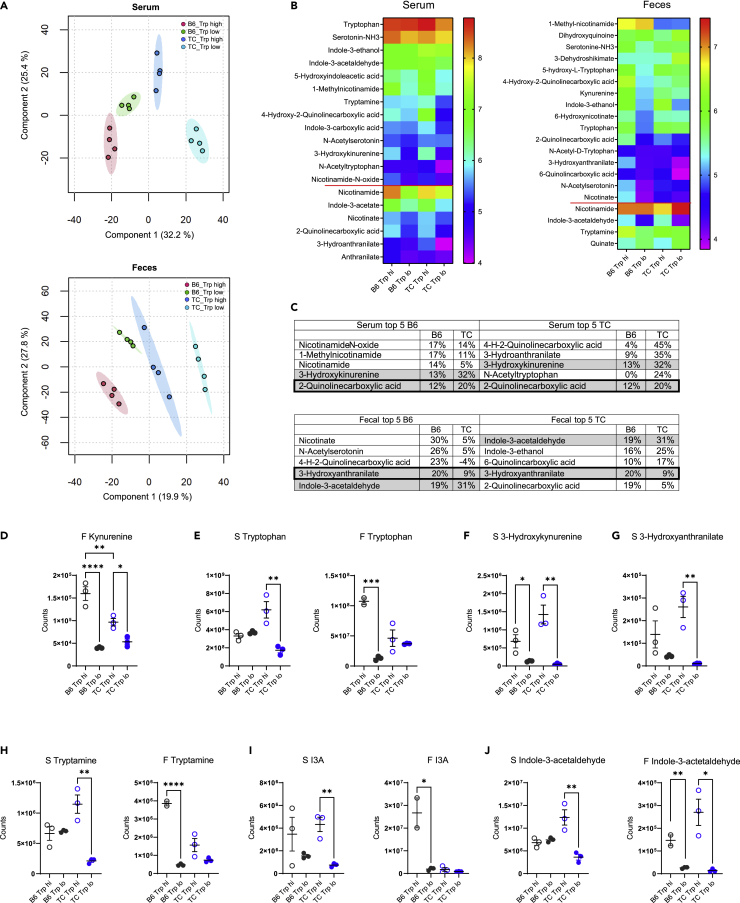
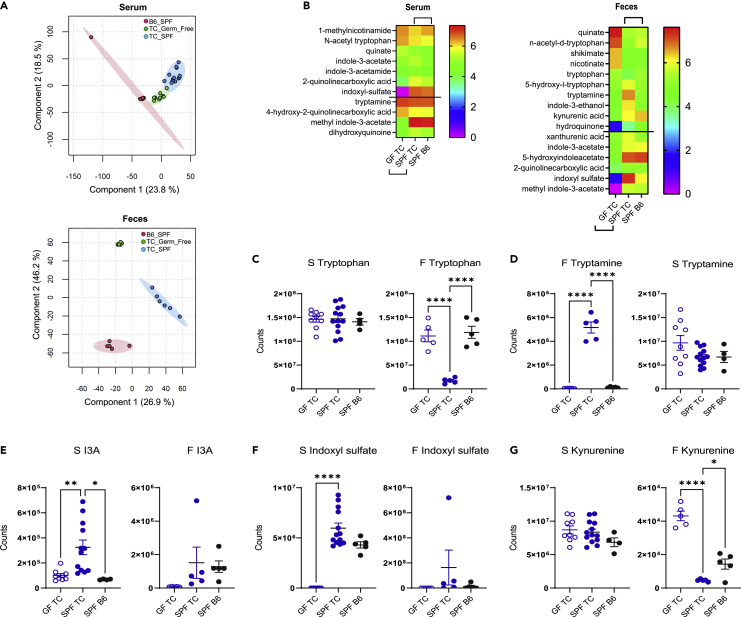
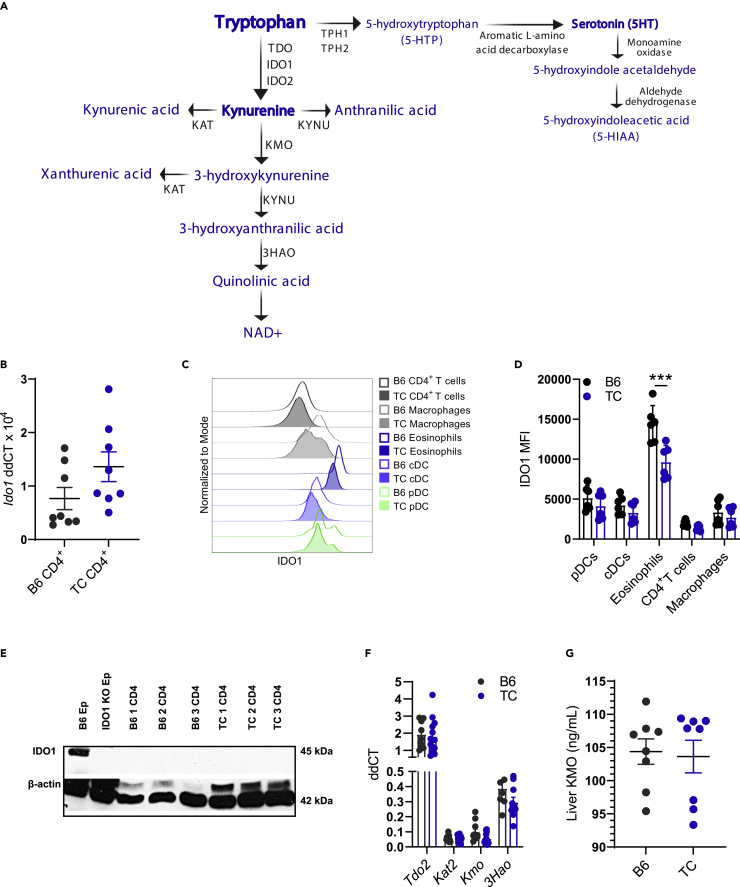
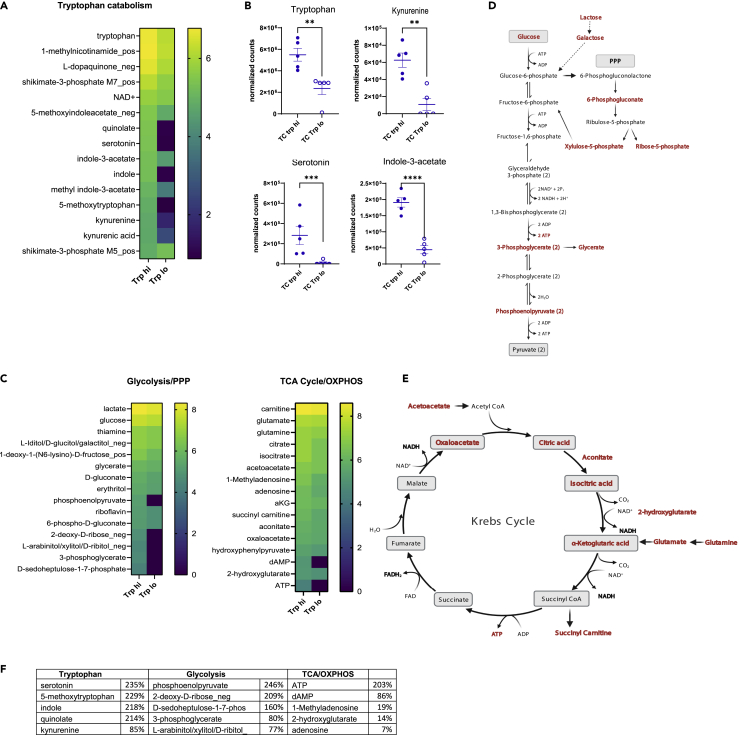
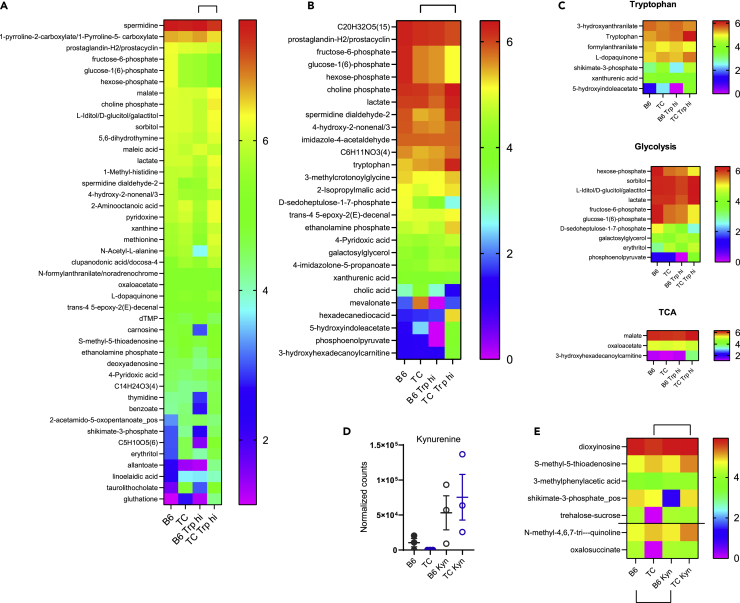
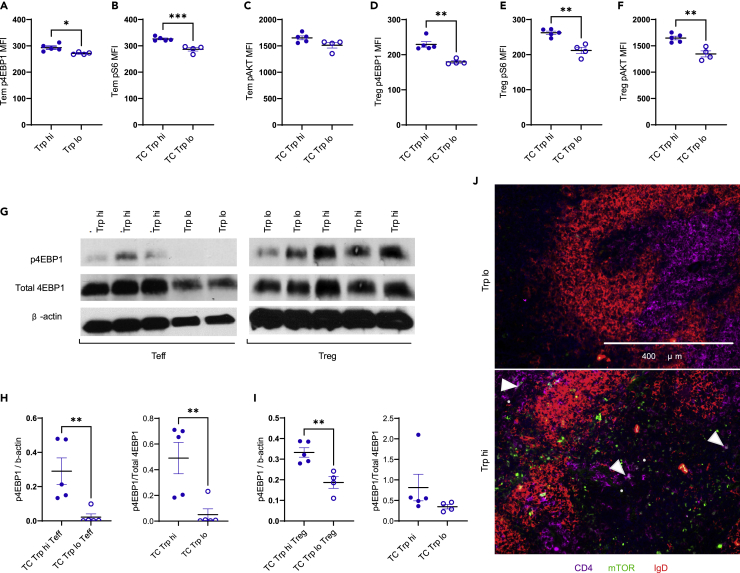
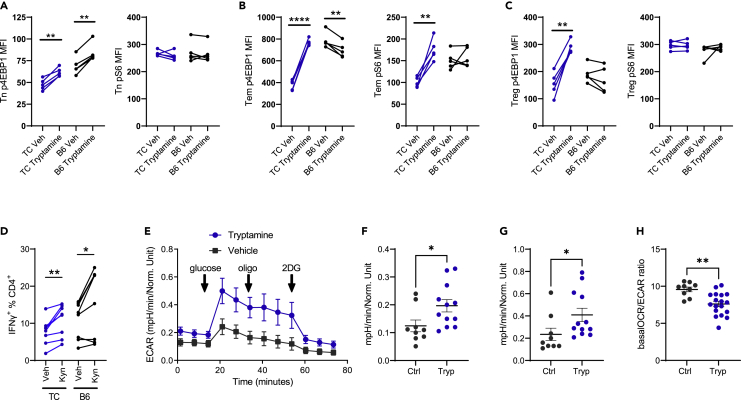
A skewed tryptophan metabolism has been reported in patients with lupus. Here, we investigated the mechanisms by which it occurs in lupus-susceptible mice, and how tryptophan metabolites exacerbate T cell activation. Metabolomic analyses demonstrated that tryptophan is differentially catabolized in lupus mice compared to controls and that the microbiota played a role in this skewing. There was no evidence for differential expression of tryptophan catabolic enzymes in lupus mice, further supporting a major contribution of the microbiota to skewing. However, isolated lupus T cells processed tryptophan differently, suggesting a contribution of T cell intrinsic factors. Functionally, tryptophan and its microbial product tryptamine increased T cell metabolism and mTOR activation, while kynurenine promoted interferon gamma production, all of which have been associated with lupus. These results showed that a combination of microbial and T cell intrinsic factors promotes the production of tryptophan metabolites that enhance inflammatory phenotypes in lupus T cells.
Keywords: Biological sciences; Cell biology; Human metabolism; Immunology.
© 2022 The Author(s).
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Åkesson K., Pettersson S., Ståhl S., Surowiec I., Hedenström M., Eketjäll S., Trygg J., Jakobsson P.J., Gunnarsson I., Svenungsson E., Idborg H. Kynurenine pathway is altered in patients with SLE and associated with severe fatigue. Lupus Sci. Med. 2018;5:e000254. doi: 10.1136/lupus-2017-000254. - DOI - PMC - PubMed
-
- Azzouz D., Omarbekova A., Heguy A., Schwudke D., Gisch N., Rovin B.H., Caricchio R., Buyon J.P., Alekseyenko A.V., Silverman G.J. Lupus nephritis is linked to disease-activity associated expansions and immunity to a gut commensal. Ann. Rheum. Dis. 2019;78:947–956. doi: 10.1136/annrheumdis-2018-214856. - DOI - PMC - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
