

Iodate Reduction by Shewanella oneidensis Requires Genes Encoding an Extracellular Dimethylsulfoxide Reductase
- PMID: 35495678
- PMCID: PMC9048795
- DOI: 10.3389/fmicb.2022.852942
Iodate Reduction by Shewanella oneidensis Requires Genes Encoding an Extracellular Dimethylsulfoxide Reductase
Abstract
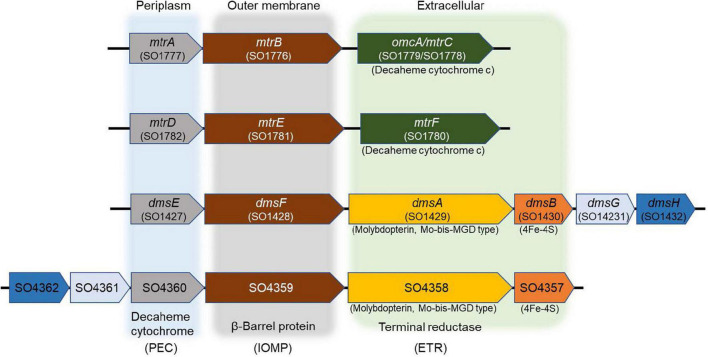
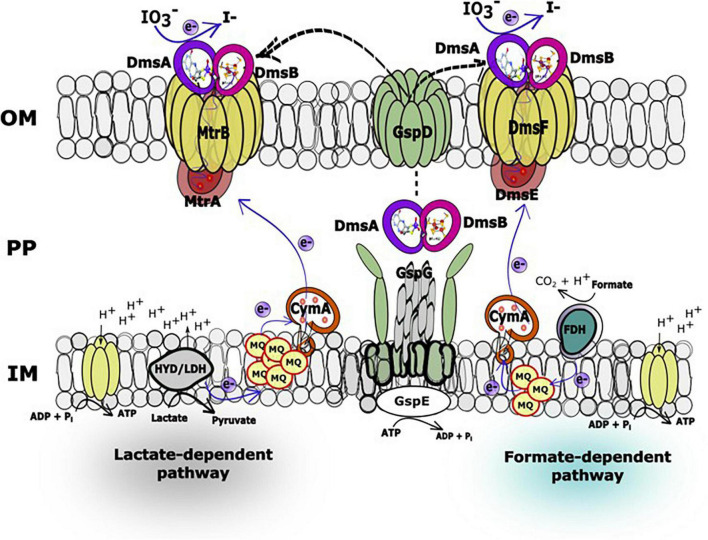
Microbial iodate (IO3 -) reduction is a major component of the iodine biogeochemical reaction network in anaerobic marine basins and radioactive iodine-contaminated subsurface environments. Alternative iodine remediation technologies include microbial reduction of IO3 - to iodide (I-) and microbial methylation of I- to volatile gases. The metal reduction pathway is required for anaerobic IO3 - respiration by the gammaproteobacterium Shewanella oneidensis. However, the terminal IO3 - reductase and additional enzymes involved in the S. oneidensis IO3 - electron transport chain have not yet been identified. In this study, gene deletion mutants deficient in four extracellular electron conduits (EECs; ΔmtrA, ΔmtrA-ΔmtrDEF, ΔmtrA-ΔdmsEF, ΔmtrA-ΔSO4360) and DMSO reductase (ΔdmsB) of S. oneidensis were constructed and examined for anaerobic IO3 - reduction activity with either 20 mM lactate or formate as an electron donor. IO3 - reduction rate experiments were conducted under anaerobic conditions in defined minimal medium amended with 250 μM IO3 - as anaerobic electron acceptor. Only the ΔmtrA mutant displayed a severe deficiency in IO3 - reduction activity with lactate as the electron donor, which suggested that the EEC-associated decaheme cytochrome was required for lactate-dependent IO3 - reduction. The ΔmtrA-ΔdmsEF triple mutant displayed a severe deficiency in IO3 - reduction activity with formate as the electron donor, whereas ΔmtrA-ΔmtrDEF and ΔmtrA-ΔSO4360 retained moderate IO3 - reduction activity, which suggested that the EEC-associated dimethylsulfoxide (DMSO) reductase membrane-spanning protein DmsE, but not MtrA, was required for formate-dependent IO3 - reduction. Furthermore, gene deletion mutant ΔdmsB (deficient in the extracellular terminal DMSO reductase protein DmsB) and wild-type cells grown with tungsten replacing molybdenum (a required co-factor for DmsA catalytic activity) in defined growth medium were unable to reduce IO3 - with either lactate or formate as the electron donor, which indicated that the DmsAB complex functions as an extracellular IO3 - terminal reductase for both electron donors. Results of this study provide complementary genetic and phenotypic evidence that the extracellular DMSO reductase complex DmsAB of S. oneidensis displays broad substrate specificity and reduces IO3 - as an alternate terminal electron acceptor.
Keywords: DMSO reductase; Shewanella oneidensis; anaerobic respiration; bioremediation; formate metabolism; iodate reduction; molybdopterin; radioactive iodine.
Copyright © 2022 Shin, Toporek, Mok, Maekawa, Lee, Howard and DiChristina.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures



References
-
- Amachi S. (2013). Iodine geochemistry and microbes: bacterial volatilization, accumulation, oxidation, reduction, sorption and dehalogenation of iodine. Chikyukagaku 47 209–219.
LinkOut - more resources
Full Text Sources