

Plasticity-Related Gene 5 Is Expressed in a Late Phase of Neurodifferentiation After Neuronal Cell-Fate Determination
- PMID: 35496908
- PMCID: PMC9053830
- DOI: 10.3389/fncel.2022.797588
Plasticity-Related Gene 5 Is Expressed in a Late Phase of Neurodifferentiation After Neuronal Cell-Fate Determination
Abstract
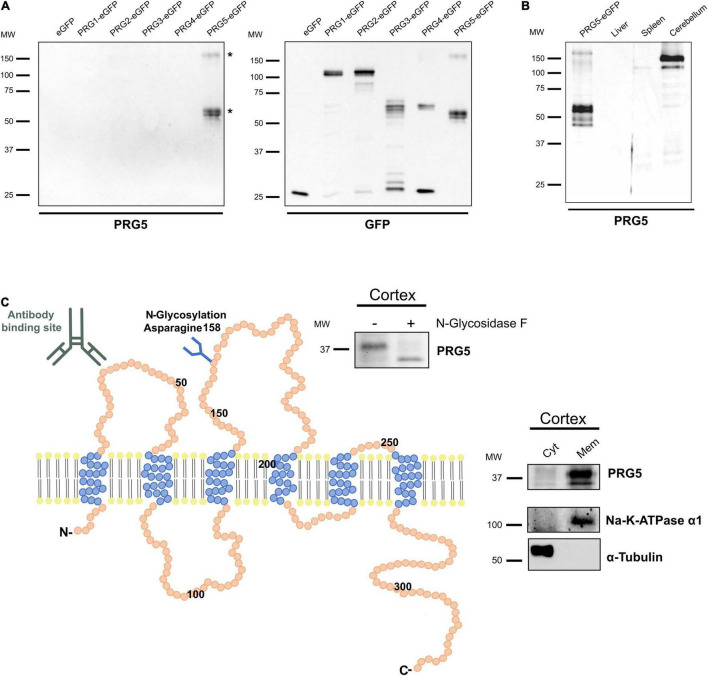
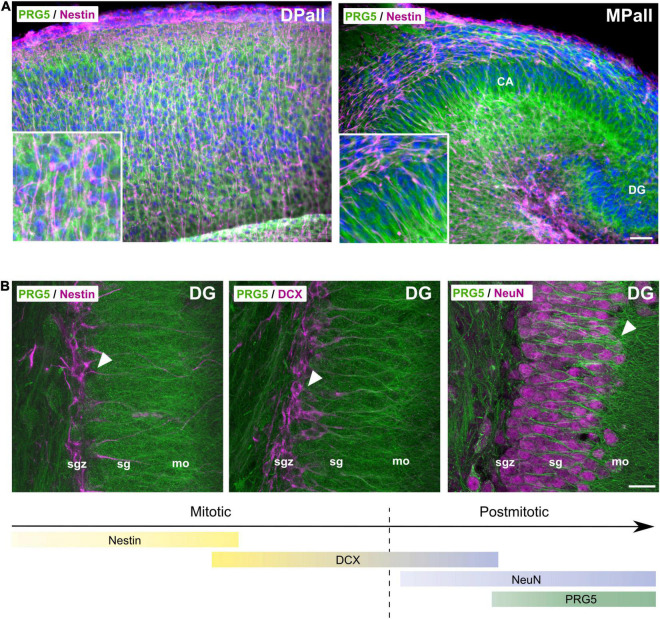
During adult neurogenesis, neuronal stem cells differentiate into mature neurons that are functionally integrated into the existing network. One hallmark during the late phase of this neurodifferentiation process is the formation of dendritic spines. These morphological specialized structures form the basis of most excitatory synapses in the brain, and are essential for neuronal communication. Additionally, dendritic spines are affected in neurological disorders, such as Alzheimer's disease or schizophrenia. However, the mechanisms underlying spinogenesis, as well as spine pathologies, are poorly understood. Plasticity-related Gene 5 (PRG5), a neuronal transmembrane protein, has previously been linked to spinogenesis in vitro. Here, we analyze endogenous expression of the PRG5 protein in different mouse brain areas, as well as on a subcellular level. We found that native PRG5 is expressed dendritically, and in high abundance in areas characterized by their regenerative capacity, such as the hippocampus and the olfactory bulb. During adult neurogenesis, PRG5 is specifically expressed in a late phase after neuronal cell-fate determination associated with dendritic spine formation. On a subcellular level, we found PRG5 not to be localized at the postsynaptic density, but at the base of the synapse. In addition, we showed that PRG5-induced formation of membrane protrusions is independent from neuronal activity, supporting a possible role in the morphology and stabilization of spines.
Keywords: PRGs; brain development; dendritic spines; neurodifferentiation; neurogenesis.
Copyright © 2022 Gross, Brandt, Vonk, Köper, Wöhlbrand, Rabus, Witt, Heep, Plösch, Hipp and Bräuer.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases